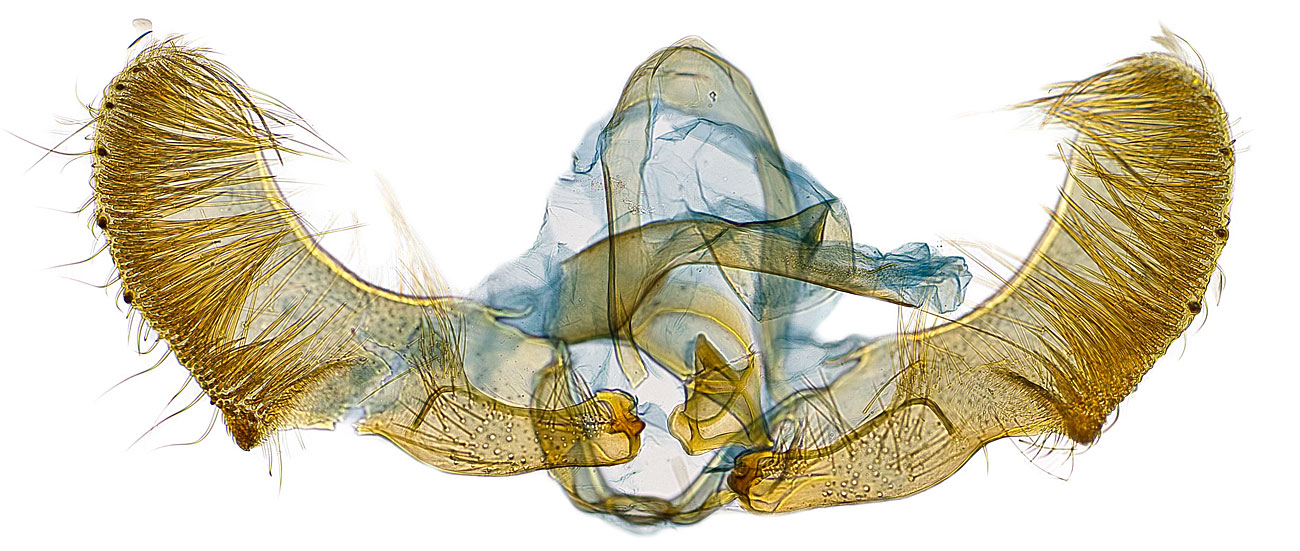
Adult Recognition

FWL: 6.0-10.5 mm
Adults are brown with indistinct reddish-brown and black markings. A conspicuous white dot is present on the distal one-third of the forewing in most individuals. Male genitalia are characterized by a dense row of ca. 130 deciduous cornuti in the vesica. Female genitalia are characterized by a cestum in the ductus bursae that is closer to the ostium than the ductus seminalis. The male antenna is flattened and notched at the base.
Males can be separated from other species of Gymnandrosoma by the flattened, notchlike basal portion of the antenna and large hairpencil on the hind tibia. Females may need to be dissected to confirm identity.
Larval Morphology
Late instar larvae are approximately 16-19 mm in length. Abdominal pinacula are large and well defined. The head is pale yellow to pale orange and the prothoracic shield is pale yellow, usually without darker mottling. Other larval characters include: distance between V setae on A9 1.5 times the distance between V setae on A8; SV pinacula on A9 unisetose; and 40-50 crochets on the abdominal prolegs.
Larvae are similar to other species in the Cryptophlebia-Ecdytolopha group, with an enlarged L-pinaculum on the prothorax that extends beneath (and usually beyond) the spiracle. Larvae of Gymnandrosoma can be separated from those of Ecdytolopha by the distance between the V setae on A9: approximately the same as the distance between Vs on A8 in Ecdytolopha and 1.2-2.0 times the distance between Vs on A8 in Gymnandrosoma.
Gymnandrosoma larvae (including G. aurantianum) are intercepted frequently on a variety of imports from Central America, South America, and the Caribbean. As most species were placed in Ecdytolopha prior to Adamski and Brown's revison (2001), interception records for Ecdytolopha could refer to species in either genus.
Biology
Gymnandrosoma aurantianum has a relatively short life cycle (36 days from egg to adult), allowing it to complete up to 10 generations per year if the appropriate host plants are available. Adults may be present year-round.
Females lay eggs on mature fruit. Larvae tunnel into fruit and consume the seeds, or they may occasionally feed on leaves and stems. Pupation occurs in the soil. Larval damage to fruit may lead to secondary infection by fungus and bacteria.
Host plants Larvae of Gymnandrosoma aurantianum can be pests on cultivated macadamia, citrus, and other tropical fruits.
| Family | Genus/species | Common name |
| Annonaceae | Annona cherimola L. X squamosa L. | |
| Euphorbiaceae | Plukenetia volubilis L. | |
| Fabaceae | Cojoba arborea (L.) Britton & Rose | |
| Fabaceae | Pithecellobium dulce (Roxb.) Benth. | monkeypod |
| Myrtaceae | Psidium guajava L. | guava |
| Oxalidaceae | Averrhoa carambola L. | carambola |
| Proteaceae | Macadamia integrifolia Maiden & Betche | macadamia nut |
| Punicaceae | Punica granatum L. | pomegranate |
| Rosaceae | Eriobotrya japonica (Thunb.) Lindl. | loquat |
| Rosaceae | Prunus persica (L.) Batsch | peach |
| Rutaceae | Citrus L. | citrus |
| Sapindaceae | Cupania vernalis A. St.-Hil. | |
| Sapindaceae | Litchi chinensis Sonn. | lychee |
| Sapindaceae | Sapindus saponaria L. | wingleaf soapberry |
| Stericulaceae | Theobroma cacao L. | cacao |
Distribution
Gymnandrosoma aurantianum is found in Central America, South America, and the Caribbean.
References
Adamski, D. and J. W. Brown. 2001. Systematic revision of the Ecdytolopha group of genera (Lepidoptera: Tortricidae: Grapholitini) in the New World. Entomologica Scandinavica Supplement 58. 86 pp.
Blanco-Metzler, H., A. D. Watt and D. Cosens. 2009. The effect of parasitism on the population dynamics of the macadamia nutborer Gymnandrosoma aurantianum (Lepidoptera: Tortricidae). Revista de Biologia Tropical. 57: 1245-1252.