Ferns produce single-celled, sexual reproductive structures called spores, which are borne in clusters on the plant’s leaves or fronds (Figure 1). Dispersal of the tiny spores is by air or water. Currently, there is no taxon identification key for ferns based solely on the relatively undifferentiated spores. However, tentative identifications, at least to genus, can be made when the disseminules are found still attached to the frond of the mother plant. Frond morphology, including the arrangement of spores, in ferns is diverse.
FNW ferns include Azolla pinnata, the Salvinia auriculata complex, Lygodium flexuosum and L. microphyllum. Their fact sheets provide pictures and more detail on the life cycles of these non-seed-bearing plants. For this identification tool, we chose images that an identifier would be able to see using a dissecting scope but not a microscope; consequently, spore photographs are not included in any of the fern fact sheets. Also, these taxa are not included in any of the keys, which are specifically based on fruit and seed disseminules.

Vegetative reproduction is common among plants. Algal species such as the FNW taxon Caulerpa taxifolia spread by means of detached segments of stem-like or leaf-like structures. Ferns, including all of the FNW species, often spread by similar means.
In angiosperms (fruited seed-bearing plants), vegetative reproduction typically results from the dispersal of an underground (e.g., rhizome) or above ground (e.g., stolon) portion of stem that includes at least one node. It is from the nodes that new roots and shoots are able to develop once the stem fragment is in a favorable environment for growth. In some cases, these modified stems have evolved into organs specialized for dormancy during unfavorable conditions. Examples include the fleshy tubers that form from rhizomes and stolons as well as bulbs and turions, which are modifications of a stem’s apex. Root tissue alone cannot produce a new plant. However, various underground stem types resemble root structures and these are often referred to as “roots” in the literature.
Taxon identification keys based on vegetative reproductive structures alone have not been developed, and probably will not be, since these structures lack sufficient character variation. More likely, DNA analysis will be used to determine taxon identification based on tissue from these structures in the future.
Many angiosperms are capable of vegetative reproduction. It is especially common in the FNW aquatic families Hydrocharitaceae and Pontederiaceae. The FNW species that disperse themselves more or less exclusively by vegetative means, and therefore, like the ferns, are not included in any of the keys based on fruit and seed disseminules, include the alga Caulerpa taxifolia and the angiosperms Hydrilla verticillata (Figures 2, 3), Lagarosiphon major, and Opuntia aurantiaca. The fact sheet for each taxon includes additional details on that taxon’s vegetative reproductive capabilities.


For angiosperms, the primary reproductive structure is the seed, which is encased in and protected by its outer coat (testa) and to varying extents by fruit tissues (pericarp). Seed and pericarp modifications primarily function to aid in dispersal of the disseminule — by providing wings or tufts of hair for sailing through the air, burs and barbed awns for attachment to animal fur, bladders for water dispersal, color and sugars for the attraction of herbivores who deposit the undigested seeds with their excrement, or explosive capsules which send seeds flying.
Since evolution serves to perfect the disseminule’s method of dispersal and not to enable us to better differentiate between seed tissue and fruit tissue, the boundaries between these tissues are sometimes hard to detect. This is particularly the case for single-seeded, dry, indehiscent (i.e., non-opening) fruits such as the grass caryopsis (discussed under Poaceae Disseminules below) or the achene, which is found in various plant families. These fruits are often casually referred to as “seeds” or “seed units” because of the overall resemblance of the unit to a true seed and because of the difficulty involved in trying to separate the seed from the fruit. The true nature of these fruits is often given away by the presence of accessory structures designed to facilitate dispersal (e.g., the tuft of hair attached to the floral end of some achenes, called cypselas, of the Asteraceae). Even though some of these structures, e.g., wings, can occur on seeds, they are more frequently found on fruits that function as disseminules (Figure 4).

A disseminule may not be strictly a seed or fruit, as seen with the legumes or pods of the Fabaceae (discussed under Fabaceae Disseminules below). A few single-seeded, dry, indehiscent fruit types have fruit tissues that are easily scraped off of the surface of the seed (e.g., the thin pericarps of utricles). In these cases, the disseminule may appear with an intact pericarp or as a naked seed.
Indehiscent but fleshy fruits, such as berries or drupes, have evolved to attract herbivores. The seeds are often swallowed whole as part of the ingested fruit and then passed onto the ground, more or less intact, with the animal's excrement. If found before ingestion but after some dessication has occurred, the disseminule may consist of seed(s) firmly encased in their shrivelled fruit coat(s) as seen on the bottom of Figure 4 (second picture from the right, which is a drupelet of Rubus moluccanus).
A few dehiscent (opening upon maturity to release the seeds) fruit types (e.g., schizocarps) split into segments (e.g., mericarps) in which some fruit tissue remains firmly attached to each seed. This fruit segment constitutes the disseminule for species with these fruit types. An example is seen in the lower left corner of Figure 4, which shows mericarps of Heracleum mantegazzianum.
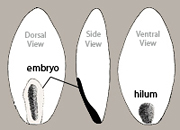
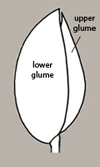
The most common dehiscent fruit type found among FNW taxa is the capsule, which is typically dry and multi-seeded. For species with capsules, the disseminule is composed of the seed alone (Figure 5). Seeds of different taxa are differentiated by several characters — including size and shape (often variable within a species), color (not always reliable), location and nature of the hilum (i.e., scar remaining where the seed had been joined to pericarp tissues via the funiculus), shape of the embryo within the testa (not always easy to tell), surface texture, and the occurrence of appendages, among others.

For FNW members of the Poaceae (grass family), a disseminule may be any of the following:
To understand these disseminules, study Figure 6 and read on.
Different disseminule types are created among grasses because species differ with respect to the location(s) of their natural disarticulation zones within the inflorescence. These zones can be above the glumes and between florets, below the glumes and between spikelets, or even lower on the floral axis, to produce clusters of spikelets.
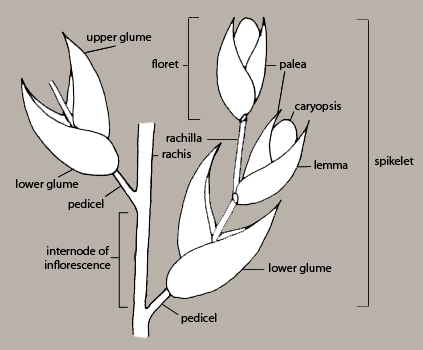
Grass species typically produce a single type of disseminule, this usually being a floret or spikelet. However, multiple types of disseminules are produced by some taxa. For the purposes of this key, it is not necessary to know in advance all of the types of disseminules that a taxon can produce. This information is in the fact sheets. Where there are normally multiple disseminule types for a taxon, separate identification entities have been constructed in the key for caryopses, florets and spikelets or clusters of spikelets as appropriate.
The caryopsis is commonly a dry, indehiscent, one-seeded fruit in which the seed is fused to the pericarp in the hilar region only. A naked caryopsis is easy to recognize. The pericarp of this small structure is often translucent, revealing the hilum at the base of one side of the fruit and the peripheral embryo (contiguous with the testa) at the base of the other side of the fruit (Figure 7). In cases where the pericarp is not translucent, the side view outline of the fruit and/or the texture of the pericarp typically reveal the position of the embryo (Figures 8, 9).

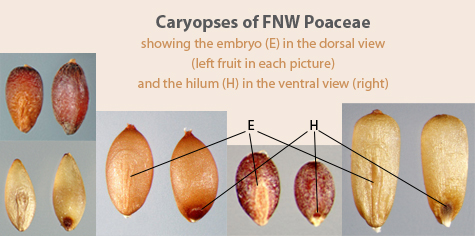
A fertile floret is not much more difficult to identify as it is merely the caryopsis tightly enclosed by 2 opposing bracts (Figure 10). The bract higher up on the axis is the palea (missing in some species where the spikelet, not the floret, is the disseminule) and is often at least partially enveloped by the lower bract, which is a fertile lemma. (Sterile lemmas, those not enclosing a caryopsis, are found in the spikelets of some FNW grass species, sometimes with an associated palea. These are known as sterile florets.)
Sometimes a floret may be mistaken for a seed or fruit of a species that is not a grass. However, under magnification, the two edges of the lemma, where it overlaps the palea, can be seen (Figure 11). More rarely only one edge is seen in species where the lemma completely envelops the palea. In either case, no other FNW fruit or seed disseminule has this characteristic.
In some grasses, a piece of the axis (called the rachilla) to which successive florets are attached may remain attached to the base of the floret (Figures 6, 12)
Another common accessory structure is the awn, which is a long, narrow bristle attached to the dorsal sides of glumes or fertile lemmas in some species (Figure 13).
Since awns are typically long relative to the size of the floret or spikelet, they are easily broken off. Sometimes dissection to remove the outer bracts and expose the lemma will reveal that a small piece of a broken awn is still attached to the lemma.
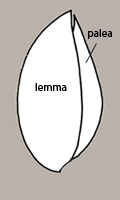



What constitutes a spikelet can be difficult to determine. It often consists of 4 or more bracts surrounding a single caryopsis, which here we refer to as a reduced spikelet. The outermost pair of bracts are called glumes — the one positioned lowest on the floral axis is called the lower glume, the other is the upper glume. The glumes sometimes entirely enclose the inner bracts and caryopsis and therefore the structure resembles a floret (Figure 14). However, the inner bracts can be revealed by teasing open the glumes at the apex of the structure when the glumes are relatively rigid or by scraping them off of the inner bracts when the glumes are more papery and fragile.
A spikelet can also be a compound structure in which 2 or more caryopses are enclosed by 2+ bracts each and then these are subtended by 1 or 2 glumes (Figures 6, 15). Or, some species shed clusters of spikelets
For some FNW grasses, spikelets or clusters of spikelets may be subtended by a leaf sheath, a whorl of hairs or an involucre (or fascicle) of bristles (Figures 16, 17).
Complicating the identification process of spikelets is that accessory structures such as easily recognized sterile florets (not those that are the lemma and palea or lemma alone appearing merely as additional bracts in the disseminule), pedicels (i.e., spikelet-bearing stalks), or parts of the inflorescence stem can also be attached to the disseminule (Figure 18).




For FNW members of the Fabaceae, a disseminule may be any of the following:
Although there is a wide range of Fabaceae disseminule types found in nature, in practice, individuals (e.g., inspectors) on the look-out for noxious weeds are most likely to encounter whole or nearly whole fruits or naked seeds, these usually being purposely transported by mail or person for medicinal uses. So, here and in the keys, we will focus on these types of disseminules.
The typical fruit of the Fabaceae is a dry, multi-seeded, elongated fruit that may be compressed or cylindrical and has two (typically visible) sutures, usually along its flattened edges (Figure 19).
Some FNW Fabaceae deviate from this standard by having fruits that are relatively short and contain few seeds, are extremely twisted or coiled, or are covered with bristles (Figure 20).
The transverse breakage of the fruits that produces single-seeded fruit segments (Figures 21, 22) is typical in Prosopis (fruit is often called a loment) and Mimosa (fruit is called a craspedium), and sometimes occurs in deeply segmented (moniliform) fruits of Acacia nilotica. The fruits of Acacia nilotica are otherwise indehiscent, while Galega officinalis is the only FNW species with a legume that dehisces along its sutures.
In general, fruit characters are more reliable than seed characters for identifying species of Prosopis and Mimosa. However, pods can change color (often darkening) and appearance (going from pubescent to glabrous) as they ripen and age. Although most of the images in the key are of more or less mature fruits, these images may represent only one developmental stage, and so the within -species variability of pod morphology is not always represented. Fruit characters that are particularly reliable for distinguishing species of Prosopis include pod proportions, number of coils, and degree of flattening. The Mimosa species have distinctly bristly pericarps.
In contrast to Prosopis, fruit characters are more variable than seed characters in Acacia nilotica. In fact, the eight infraspecific taxa of A. nilotica are defined largely by their different fruit characters, especially as they relate to pubescence and degree of segmentation of the pods between seeds.




Seeds of Prosopis and Mimosa darken with age, so seed color is not a good discriminating character. Seeds of these genera often look very similar (Figure 23) and within-species morphological variability may not be represented in the key or fact sheets, especially since relatively small numbers of seeds were available for examination for some species. In contrast, seeds of Acacia nilotica and Galega officinalis are easily recognized by their pleurogram and lens shape in the case of the former and seed shape and lack of pleurogram in the latter (Figure 23).
Useful seed characters include the shape of the pleurogram, a U- or O-shaped line occurring on both seed faces and resulting from modification of the face of the ovule during seed development (Figure 24). When present (it does not occur in Galega officinalis), it is usually visible but sometimes faint. The area within the pleurogram is typically about one third the width of the seed. Pleurograms may be broadly open to nearly closed in Prosopis and Mimosa to closed (or O-shaped) in Acacia nilotica (Figure 25). See Figure 23 for pleurogram variation in Prosopis.
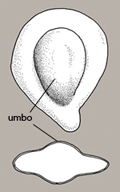
The umbo is a rounded protuberance on both faces of some of the FNW Prosopis species (Figure 26).
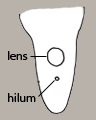
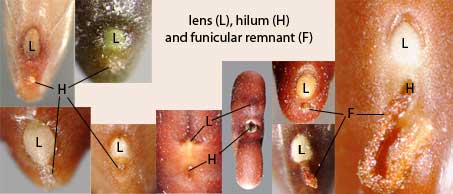
The scar on a seed where the funiculus attached the seed to the pericarp tissues is called the hilum. The lens (Figure 27) is a mound, pad, or area of tissue situated near the hilum that acts as a site of initial water entry and regulation by the otherwise relatively impermeable seed. It is often more visible than the hilum and therefore mistaken for that structure. If a funiculus remnant adheres to the hilum, which is not unusual, then the distinction between the two structures is clear. Aiding in the identification of the lens is the fact that it is typically larger than the hilum, is often whitish or yellowish, and may be raised, flush or recessed. See examples below in Figure 28.


Authors: J.L. Scher, D. S. Walters, and A.J. Redford
Edition 2.2 - April 2015; Nomenclature last updated Feb 2026
Lucid mobile app for Android and iOS | tool images at ITP Node
idtools.org
