|
|
CAPS Primary Target - AdultPort Interception Target - Larva |
Epiphyas postvittana (Walker) (Tortricidae: Tortricinae: Archipini) Common names: light brown apple moth (LBAM) Synonyms: basialbana (Teras), consociana (Pandemis), dissipata (Tortrix), foedana (Dichelia), oenopa (Tortrix), phaeosticha (Tortrix), pyrrhula (Tortrix), retractana (Dichelia), reversana (Dichelia), scitulana (Teras), secretana (Teras), stipularis (Tortrix), vicariana (Dichelia), vicaureana (Dichelia) |
|
|
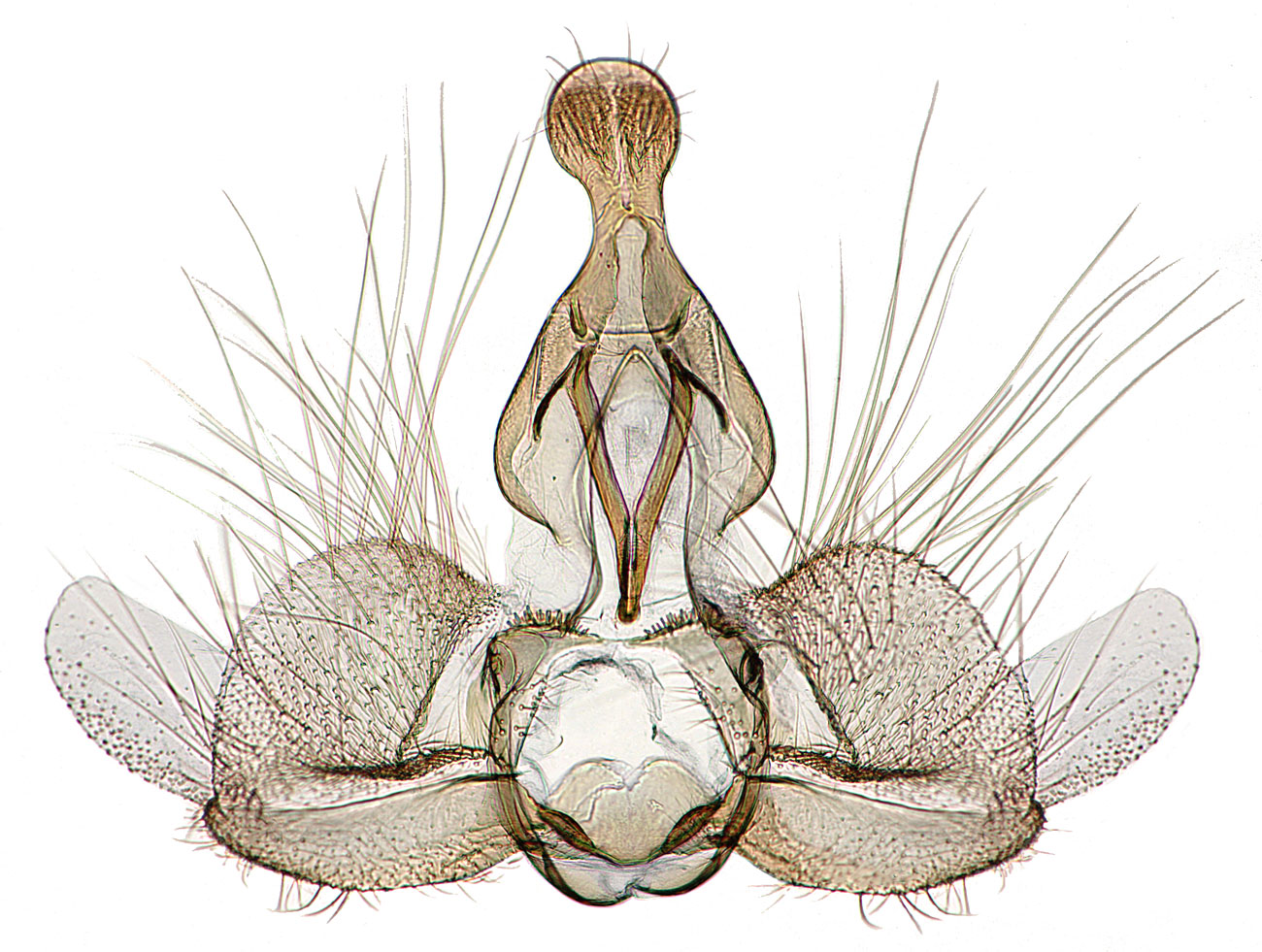
Adult Recognition

FWL: 5.0-11.5mm (male); 5.5-12.5mm (female) Forewings of both sexes are light brown to pale yellow with brown to dark-brown markings. Males are more variable than females, although in most males the basal half of the forewing is lightly marked, the median fascia is well defined, and there is a dark mark on the costa distal to the median fascia. In California, males tend to be of three phenotypes; the form with solid dark markings on the distal half of the forewing is the most uncommon. Males have a forewing costal fold. The female forewing color is more uniform, with a poorly defined median fascia and an overall mottled or speckled appearance. Most females have a dark mark on the dorsum of each forewing and two dark spots on the posterior of the thorax. The hindwing in both males and females is mottled with dark scales, although this pattern is usually more evident in females. Male genitalia are distinctive, and examination of these structures is essential for reliable identification. Males have a combination of the following characters: spatulate uncus; reduced socii; short valva with a broad sacculus; membranous lobe on the apex of the valve (the most diagnostic feature); and an aedeagus with 2-4 deciduous cornuti. Female genitalia are typical of many Archipini and females may be difficult to verify based on dissection alone. Females possess a combination of the following characters: simple sterigma; long, straight ductus bursae which is 2/3 or more the length of the abdomen; and corpus bursae with a single, hook-shaped signum. Adults are similar to other species of Epiphyas as well as many Nearctic Archipini. A dissection can be used to confirm identity. Male E. postvittana have a large membranous lobe on the apex of the valva that is not present in most Nearctic tortricids. A membranous lobe is present in many Clepsis (C. fucana, C. peritana, and C. virescana), but the lobe in E. postvittana is much larger and forms a conspicuous notch that is not present in species like C. peritana.
Larval Morphology
Larvae are generally yellowish green but color may vary with instar and host. Early and mid-instar larvae range from translucent to opaque reddish brown. The head of all instars is pale brown and the prothoracic shield is approximately the same color as the rest of the body. The head, prothoracic shield, and legs of mid- to late instars are not dark and do not have any dark or contrasting markings. The anal shield is pale brownish green and the anal comb is well developed with 7-9 teeth. First instar larvae are approximately 1.6 mm long, while last instar larvae are 10-20 mm in length. Chaetotaxy is typical of most Tortricidae, with a trisetose L group on the prothorax; L1 and L2 on a common pinaculum below the spiracle on A1-7; L1 anterad of the spiracle on A8; and D2s on a common "saddle" pinaculum on A9.Typical of most Archipini, the small SD2 pinaculum is fused with the anterior edge of the large SD1 pinaculum on A1-7; D1 and SD2 are on separate pinacula on A9; the L group is trisetose with all setae on the same pinaculum on A9; the SV group on A1,2,7,8,9 is 3:3:3:2:2; V1s on A9 are nearly the same distance or only slightly further apart than those on A7 and A8. On the head capsule, AF2 and P1 are approximately the same distance apart as P1 and P2. Larvae are similar to those of several other common California tortricids, including Argyrotaenia franciscana and Platynota stultana. Molecular diagnostics may be necessary to positively identify E. postvittana larvae, especially early instars.
Biology
Epiphyas postvittana completes 2-4 generations annually over much of its range, depending on temperature and latitude. Populations in California appear to undergo at least four generations and adults are active throughout the year. The upper and lower temperature thresholds for development in laboratory studies are 7.5 and 31°C; 20°C is the optimum for development, leading to a life cycle of 25 days. Females deposit eggs in masses on smooth surfaces of host plant foliage, including leaves, stems, and fruit. Egg masses vary from 4 to 150 individual eggs. Larvae pass through five or six instars; they can overwinter, but development during cooler months is slower. Early instar larvae feed on the undersides of leaves within a silk chamber. Later instars may fold individual leaves, create a nest of several leaves webbed together, or web leaves to fruit and feed on the surface of the fruit. Larvae on deciduous trees and shrubs feed as long as leaves remain on the host plant and then drop to the ground, where they may feed on understory vegetation or survive in leaf litter. Pupation occurs in the larval nest, and adults eclose in approximately 10 days at 20°C. Host plants Epiphyas postvittana is a polyphagous pest on pome and stone fruits and other horticultural crops. It has been recorded from more than 500 plant species in 121 families and 363 genera, although larvae prefer herbaceous plants over woody ones. Larvae feed on the leaves, buds, flowers, and fruits of their hosts, but the majority of economic damage is caused by fruit injury. Larvae feed on the surface of fruits under webbed leaves, causing scarring as well as providing a site for rot or infection. In Australia and New Zealand larvae can be a pest on pome and stone fruits as well as other horticultural crops. If left uncontrolled, larval damage to fruit crops in Australia and New Zealand during years of high population densities has been as high as 70% (with a typical range of 5-70%). The following is a partial host list. | Family | Genus/species | Common name | | Actinidaceae | Actinidia Lindl. | kiwi | | Aizoaceae | Mesembryanthemum L. | iceplant | | Amaranthaceae | Amaranthus L. | pigweed | | Anacardiaceae | Mangifera indica L. | mango | | Apiaceae | Daucus L. | wild carrot | | Apiaceae | Petroselinum J. Hill | parsley | | Apiaceae | Platysace Bunge | | | Apocynaceae | Vinca L. | periwinkle | | Araliaceae | Hedera L. | ivy | | Asteraceae | Arctotheca calendula (L.) Levyns | capeweed | | Asteraceae | Artemisia L. | sagebrush | | Asteraceae | Aster L. | aster | | Asteraceae | Baccharis L. | baccharis | | Asteraceae | Calendula L. | marigold | | Asteraceae | Chrysanthemum L. | daisy | | Asteraceae | Dahlia Cav. | dahlia | | Asteraceae | Erigeron L. | fleabane | | Asteraceae | Gerbera J. F. Gmel. | Transvaal daisy | | Asteraceae | Helichrysum Mill. | strawflower | | Asteraceae | Senecio L. | ragwort | | Asteraceae | Tithonia Desf. ex Juss. | tithonia | | Betulaceae | Alnus glutinosa (L.) Gaertn. | European alder | | Bignoniaceae | Campsis Lour. | trumpet-vine | | Brassicaceae | Brassica L. | mustard | | Brassicaceae | Raphanus L. | radish | | Brassicaceae | Sisymbrium L. | hedgemustard | | Buddlejaceae | Buddleja L. | butterflybush | | Cannabaceae | Humulus lupulus L. | common hop | | Caprifoliaceae | Lonicera L. | honeysuckle | | Caprifoliaceae | Viburnum L. | viburnum | | Celastraceae | Euonymus L. | spindletree | | Chenopodiaceae | Chenopodium L. | goosefoot | | Clusiaceae | Hypericum perforatum L. | common St. Johnswort | | Cupressaceae | Cupressus L. | cypress | | Ebenaceae | Diospyros kaki L. f. | Japanese persimmon | | Ebenaceae | Diospyros L. | diospyros | | Epacridaceae | Monotoca R. Br. | | | Ericaceae | Arbutus L. | madrone | | Ericaceae | Vaccinium L. | blueberry | | Escalloniaceae | Escallonia Mutis ex L.f. | escallonia | | Euphorbiaceae | Breynia J.R. Forst. & G. Forst. | breynia | | Euphorbiaceae | Phyllanthus L. | leafflower | | Fabaceae | Acacia Mill. | acacia | | Fabaceae | Cassia L. | cassia | | Fabaceae | Cytisus scoparius (L.) Link | Scotch broom | | Fabaceae | Genista L. | broom | | Fabaceae | Hardenbergia Benth. | | | Fabaceae | Lathyrus L. | pea | | Fabaceae | Lupinus L. | lupine | | Fabaceae | Medicago sativa L. | alfalfa | | Fabaceae | Trifolium L. | clover | | Fabaceae | Ulex europaeus L. | common gorse | | Fabaceae | Vicia faba L. | horsebean | | Fagaceae | Quercus L. | oak | | Geraniaceae | Pelargonium L'Her. ex Aiton | geranium | | Grossulariaceae | Ribes L. | currant | | Hydrangeaceae | Philadelphus L. | mock orange | | Iridaceae | Crocosmia Planch. | crocosmia | | Juglandaceae | Juglans L. | walnut | | Juncaginaceae | Triglochin L. | arrowgrass | | Lamiaceae | Lavandula L. | lavender | | Lamiaceae | Mentha L. | mint | | Lamiaceae | Salvia L. | sage | | Lauraceae | Persea americana Mill. | avocado | | Loganaceae | Gelsemium Juss. | trumpetflower | | Magnoliaceae | Michelia L. | michelia | | Malvaceae | Sida L. | fanpetals | | Myoporaceae | Myoporum Sol. ex G. Forst. | myoporum | | Myrtaceae | Acca sellowiana (O. Berg.) Burret | feijoa | | Myrtaceae | Astartea DC. | | | Myrtaceae | Callistemon R. Br. | bottlebrush | | Myrtaceae | Eucalyptus L'Her. | gum | | Myrtaceae | Leptospermum J.R. Forst. & G. Forst. | teatree | | Myrtaceae | Melaleuca L. | bottlebrush | | Oleaceae | Forsythia Vahl | fosythia | | Oleaceae | Jasminum L. | jasmine | | Oleaceae | Ligustrum L. | privet | | Oxalidaceae | Oxalis L. | woodsorrel | | Pinaceae | Pinus L. | pine | | Pittosporaceae | Billardiera Sm. | | | Pittosporaceae | Bursaria Cav. | | | Pittosporaceae | Pittosporum Banks ex Sol. | cheesewood | | Plantaginaceae | Plantago lanceolata L. | narrowleaf plantain | | Polygonaceae | Polygala L. | milkwort | | Polygonaceae | Polygonum L. | knotweed | | Polygonaceae | Rumex L. | dock | | Proteaceae | Grevillea R. Br. ex Knight | grevillea | | Proteaceae | Leucadendron L. | | | Proteaceae | Macadamia integrifolia Maiden & Betche | macadamia nut | | Proteaceae | Persoonia Sm. | | | Pteridaceae | Adiantum L. | maidenhair fern | | Pteridaceae | Pteris L. | brake fern | | Ranunculaceae | Aquilegia L. | columbine | | Ranunculaceae | Clematis L. | leather flower | | Ranunculaceae | Ranunculus L. | buttercup | | Resedaceae | Reseda L. | mignonette | | Rhamnaceae | Ceanothus L. | ceanothus | | Rosaceae | Cotoneaster Medik. | cotoneaster | | Rosaceae | Crataegus L. | hawthorn | | Rosaceae | Cydonia oblonga Mill. | quince | | Rosaceae | Eriobotrya japonica (Thunb.) Lindl. | loquat | | Rosaceae | Fragaria L. | strawberry | | Rosaceae | Malus Mill. | apple | | Rosaceae | Photinia Lindl. | chokeberry | | Rosaceae | Prunus armeniaca L. | apricot | | Rosaceae | Prunus persica (L.) Batsch | peach | | Rosaceae | Pyracantha M. Roem. | firethorn | | Rosaceae | Pyrus L. | pear | | Rosaceae | Rosa L. | rose | | Rosaceae | Rubus L. | [various] | | Rutaceae | Boronia Sm. | boronia | | Rutaceae | Choisya Kunth | Mexican orange | | Rutaceae | Citrus L. | citrus | | Rutaceae | Correa Andrews | Australian fuschia | | Rutaceae | Eriostemon Sm. | | | Rutaceae | Fortunella Swingle | kumquat | | Salicaceae | Populus L. | cottonwood | | Salicaceae | Salix L. | willow | | Sapindaceae | Dodonaea Mill. | dodonaea | | Sapindaceae | Litchi chinensis Sonn. | lychee | | Scrophulariaceae | Hebe Comm. ex Juss. | hebe | | Smilacaceae | Smilax L. | greenbrier | | Solanaceae | Datura L. | jimsonweed | | Solanaceae | Solanum lycopersicum L. var. lycopersicum | garden tomato | | Solanaceae | Solanum tuberosum L. | Irish potato | | Theaceae | Camellia japonica L. | | | Ulmaceae | Trema Lour. | trema | | Urticaceae | Urtica L. | nettle | | Valerianaceae | Centranthus Neck. ex Lam. & DC. | fox-brush | | Verbenaceae | Clerodendron Burm. | | | Vitaceae | Parthenocissus Planch. | creeper | | Vitaceae | Vitis L. | grape |
Distribution
A native of Australia, E. postvittana occurs in all apple-growing areas of that country. It has been introduced into Tasmania, New Zealand and Hawaii. In Europe, it has been present in the United Kingdom since the mid-1930s and is intermittently recorded from continental Europe, most recently in the Netherlands and Sweden. Epiphyas postvittana was first discovered in California in 2006. It is currently well-established and has been recorded from 23 California counties. In early 2011, a single adult was captured in Oregon, but an established population has not been found in the U.S. outside of California.
References
Barr, N. B., L. A. Ledezma, J. D. Vasquez, M. E. Epstein, P. H. Kerr, S. Kinnee, O. Sage and T. M. Gilligan. 2009. Molecular identification of the light brown apple moth (Lepidoptera: Tortricidae) in California using a polymerase chain reaction assay of the internal transcribed spacer 2 locus. Journal of Economic Entomology. 102(6): 2333-2342. Barr, N. B., L. A. Ledezma, R. E. Farris, M. E. Epstein and T. M. Gilligan. 2011. A multiplex real-time polymerase chain reaction assay to diagnose Epiphyas postvittana (Lepidoptera: Tortricidae). Journal of Economic Entomology. 104(5): 1706-1719. Brown, J. W., M. E. Epstein, T. M. Gilligan, S. Passoa and J. A. Powell. 2010. Biology, identification, and history of the light brown apple moth, Epiphyas postvittana (Walker) (Lepidoptera: Tortricidae: Archipini) in California: an example of the importance of local faunal surveys to document the establishment of exotic insects. American Entomologist. 56(1): 34-43. Danthanarayana, W. 1975. The bionomics, distribution and host range of the light brown apple moth, Epiphyas postvittana (Walk.) (Tortricidae). Australian Journal of Zoology 23: 419-437. Geier, P. and D. Briese. 1981. The light-brown apple moth, Epiphyas postvittana (Walker): a native leafroller fostered by European settlement., pp. 131-155. In R. Kitching and R. Jones [eds.], The Ecology of Pests. CSIRO, Melbourne, Australia. Gilligan, T. M. and M. E. Epstein. 2009. LBAM ID: Tools for diagnosing light brown apple moth and related western U.S. leafrollers (Tortricidae: Archipini). CDROM. Center for Plant Health Science and Technology, USDA, APHIS PPQ, Raleigh, NC. Gu, H. and W. Danthanarayana. 1992. Influence of larval rearing conditions on the body size and flight capacity of Epiphyas postvittana moths. Australian Journal of Zoology 40: 573-581. Suckling D. M., J. G. I. Khoo and D. J. Rogers. 1990. Disruption of lightbrown apple moth Epiphyas postvittana (Lepidoptera: Tortricidae) trapping in Nelson, New Zealand. Australian Journal of Zoology 38: 363-373. Suckling, D. M. and E. G. Brockerhoff. 2010. Invasion biology, ecology, and management of the light brown apple moth (Tortricidae). Annual Review of Entomology. 55: 285-306. Tooman, L. K., C. J. Rose, C. Carraher, D. M. Suckling, S. R. Paquette, L. A. Ledezma, T. M. Gilligan, M. Epstein, N. B. Barr and R. D. Newcomb. 2011. Patterns of mitochondrial haplotypes diversity in the invasive pest Epiphyas postvittana (Lepidoptera: Tortricidae). Journal of Economic Entomology. 104(3): 920-932. Wearing C. H., W. P. Thomas, J. W. Dugdale and W. Danthanarayana. 1991. Tortricid pests of pome and stonefruits, Australian and New Zealand species., pp. 453-472. In L. P. S. van der Geest and H. H. Evenhius [eds.], Tortricid Pests: Their Biology, Natural Enemies, and Control. World Crop Pests, Vol. 5. Elsevier, Amsterdam.
|
|
Tortricids of Agricultural Importance by Todd M. Gilligan and Marc E. Epstein
Interactive Keys developed in Lucid 3.5. Last updated August 2014.
|
|