Noctuoidea: Noctuidae: Heliothinae: Helicoverpa zea (Boddie)
corn earworm, tomato fruitworm, cotton bollworm
Heliothis zea, H. umbrosus, H. ochracea, H. stombleri
Prior to the mid-20th century, the name armigera was often used to refer to H. zea in the New World.
The majority of interception records (>70%) are from Mexico, with nearly half of those associated with corn (Zea mays). Larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of H. zea are highly polyphagous and interceptions have been recorded from more than 180 types of plants. Records in PestID from locations outside of the Americas or the Caribbean could represent H. zea or any of several sibling species with poorly known larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
.
| Origin | Host(s) |
|---|---|
| Dominican Republic | Capsicum |
| Ecuador | various |
| Guatemala | various |
| Mexico | Capsicum, Cicer, Lactuca, Ocimum, Phaseolus, Physalis, Zea mays |
| Nicaragua | various |
| Peru | Pisum, Zea mays |
| Trinidad and Tobago | Capsicum |
Helicoverpa zea is widely distributed throughout the Americas. It is present in North, Central, and South America, the Caribbean, and Hawaii. It has also been recorded from other Pacific Islands (populations which likely did not establish) (Hardwick 1965Hardwick 1965:
Hardwick, D. F. 1965. The corn earworm complex. Memoirs of the Entomological Society of Canada 40: 1-247.).
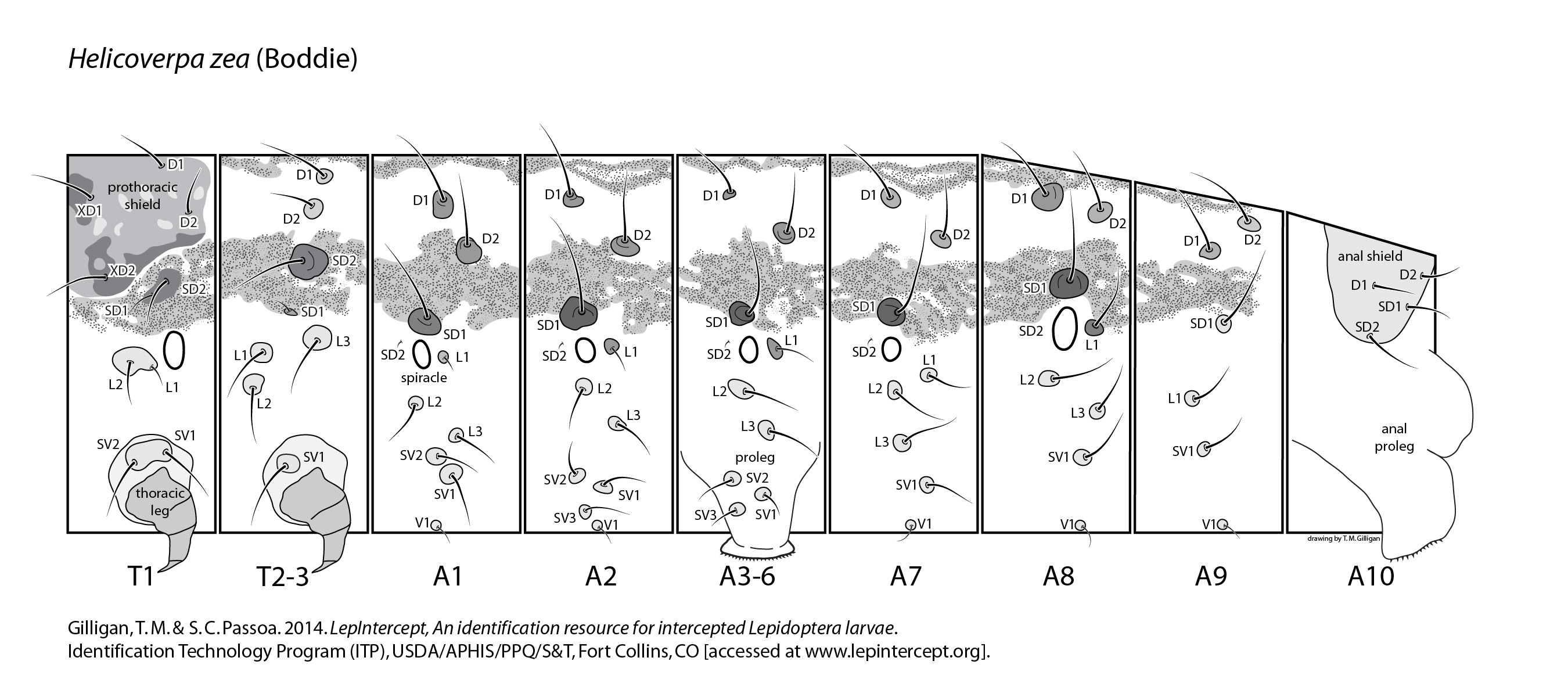
Only mid- to late instar larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
can be identified using the mandible and lack of microspinesmicrospines:
minute spines on the body, usually visible only under magnification
on the pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
; early instars should be identified only to subfamily. Although H. zea is only recorded from the New World, the presence of other Helicoverpa can make a species-level identification difficult. Consult the Detailed Information tab for information on other species present in South America. The discovery of H. armigera in Brazil, Paraguay, and Argentina makes species-level identifications from those countries impossible using only morphology.
The larvalarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of the corn earworm, Helicoverpa zea, was at least partially described by many authors including Garman (1920)Garman (1920):
Garman, H. 1920. Observations on the structure and coloration of the larval corn-ear worm (Chloridea obsoleta), the bud worm (Chloridea virescens) and a few other lepidopterous larvae. Kentucky Agricultural Experiment Station Bulletin 227: 55-84., Crumb (1956)Crumb (1956):
Crumb, S. E. 1956. The Larvae of the Phalaenidae. U. S. Department of Agriculture Technical Bulletin 1135. 356 pp., Okumura (1961)Okumura (1961):
Okumura, G. T. 1961. Identification of lepidopterous larvae attacking cotton. California Department of Agriculture Bureau of Entomology Special Publication 282. 80 pp., Peterson (1962)Peterson (1962):
Peterson, A. 1962. Larvae of insects: an introduction to Nearctic species. Part I: Lepidoptera and plant infesting Hymenoptera. Columbus, Ohio. 315 pp., Hardwick (1965)Hardwick (1965):
Hardwick, D. F. 1965. The corn earworm complex. Memoirs of the Entomological Society of Canada 40: 1-247., Neunzig (1969)Neunzig (1969):
Neunzig, H. H. 1969. The biology of the tobacco budworm and the corn earworm in North Carolina with particular reference to tobacco as a host. North Carolina Agricultural Experiment Station Technical Bulletin 196. 76 pp., Pastrana and Henandez (1979), Godfrey (1987)Godfrey (1987):
Godfrey, G. L. 1987. Noctuidae (Noctuoidea), pp. 549-578. In F. W Stehr (ed.). Immature Insects. Kendall Hunt Publishing Company. Dubuque, Iowa., and Hardwick (1996)Hardwick (1996):
Hardwick, D. F. 1996. A Monograph to the North American Heliothentinae (Lepidoptera: Noctuidae). Almonte, Ontario [privately published]. 281 pp.. Several larval color patterns were photographed by Hardwick (1996)Hardwick (1996):
Hardwick, D. F. 1996. A Monograph to the North American Heliothentinae (Lepidoptera: Noctuidae). Almonte, Ontario [privately published]. 281 pp., Cranshaw (2004)Cranshaw (2004):
Cranshaw, W. 2004. Garden Insects of North America. Princeton University Press. Princeton, New Jersey. 678 pp., and Wagner et al. (2011)Wagner et al. (2011):
Wagner, D. L., D. F. Schweitzer, J. B. Sullivan and R. C. Reardon. 2011. Owlet caterpillars of eastern North America. Princeton University Press, New Jersey. 576 pp.. Brazzel et al. (1953)Brazzel et al. (1953):
Brazzel J. R., L. D. Newsom, J. S. Roussel, C. Lincoln, F. J. Williams and G. Barnes. 1953. Bollworm and tobacco budworm as cotton pests in Louisiana and Arkansas. Louisiana Technical Bulletin 482. 47 pp. illustrated variation in the mandibles of H. zea. Early literature was reviewed Kogan et al. (1978)Kogan et al. (1978):
Kogan, J., D. K. Sell, R. E. Stinner, J. R. Bradley Jr. and M. Kogan. 1978. A Bibliography of Heliothis zea (Boddie) and H. virescens (F.) (Lepidoptera: Noctuidae). International Soybean Program series 17. Urbana, Illinois. 242 pp.. The Heliothinae guide (below) provides keys to separate H. zea from other quarantine species of Heliothinae.
Larval variation in the Chloridea/Heliothis/Helicoverpa complex can complicate identifications. Although simplified keys work for most situations in the United States, APHIS intercepts so many individuals that exceptions are common and have to be discussed.
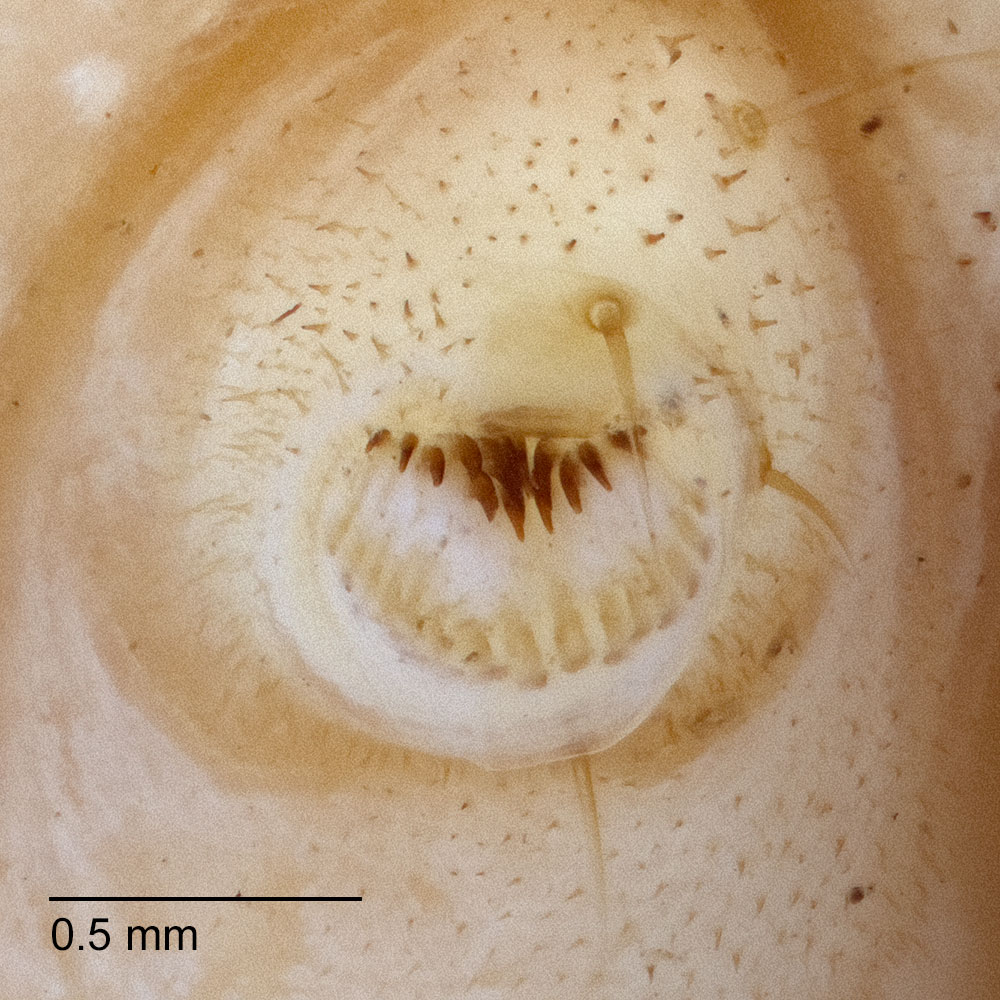
Typically, H. zea is recognized by having no retinaculumretinaculum:
a projection or toothlike structure on the oral surface of the mandible
on the mandible and a lack of microspinesmicrospines:
minute spines on the body, usually visible only under magnification
on the body pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
(e.g., Godfrey 1987Godfrey 1987:
Godfrey, G. L. 1987. Noctuidae (Noctuoidea), pp. 549-578. In F. W Stehr (ed.). Immature Insects. Kendall Hunt Publishing Company. Dubuque, Iowa.). These characters separate H. zea from most Chloridea virescens. The mandible of H. zea normally lacks a retinaculumretinaculum:
a projection or toothlike structure on the oral surface of the mandible
, although a small tooth may be present. A retinaculumretinaculum:
a projection or toothlike structure on the oral surface of the mandible
, usually well developed in C. virescens, may be reduced to a thin ridge or "scar." Each of these cases was illustrated by Brazzel et al. (1953)Brazzel et al. (1953):
Brazzel J. R., L. D. Newsom, J. S. Roussel, C. Lincoln, F. J. Williams and G. Barnes. 1953. Bollworm and tobacco budworm as cotton pests in Louisiana and Arkansas. Louisiana Technical Bulletin 482. 47 pp. and Neunzig (1969: 11). A small number of microspinesmicrospines:
minute spines on the body, usually visible only under magnification
may be found on the body pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
of H. zea in the fifth and sixth instars, but only around the edges (Brazzel et al. 1953: 16). The microspinesmicrospines:
minute spines on the body, usually visible only under magnification
of C. virescens tend to reach at least the middle of the body pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
on many segments (see C. virescens fact sheet). Boyer et al. (1977)Boyer et al. (1977):
Boyer, W. P., J. G. Burleigh and M. L. Wall. 1977. Larval characters for separating bollworm and tobacco budworm. Annals of the Entomological Society of America 70: 5-6. suggested the most accurate way to evaluation the microspine character was to use the dorsal pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
on A8.
Typically, Heliothis phloxiphaga has conical pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
all over the body. Conical pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
in H. zea, if present, are only on A1, A2, and A8. Some specimens of H. phloxiphaga have flat pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
(Hardwick 1996: N7, N8); these can be confused with H. zea. Unlike H. zea, H. phloxiphaga sometimes has dark arcs on the head (Crumb 1926Crumb 1926:
Crumb, S. E. 1926. The Nearctic budworms of the lepidopterous genus Heliothis. Proceedings of the United States National Museum 68: 1-8.) or pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
ringed with white (Lange and Michelbacher 1937Lange and Michelbacher 1937:
Lange, W. H. and A. E. Michelbacher. 1937. Two closely related species of Heliothis found in tomato fields of central California. The Bulletin of the Department of Agriculture, State of California 26: 320-325.).
Normally, H. zea does not feed on Physalis (Godfrey 1987Godfrey 1987:
Godfrey, G. L. 1987. Noctuidae (Noctuoidea), pp. 549-578. In F. W Stehr (ed.). Immature Insects. Kendall Hunt Publishing Company. Dubuque, Iowa.), while Chloridea subflexa is a Physalis specialist. However, Robinson et al. (2002)Robinson et al. (2002):
Robinson G. S., P. R. Ackery, I. J. Kitching, G. W. Beccaloni and L. M. Hernandez. 2002. Hostplants of the moth and butterfly caterpillars of America north of Mexico. Memoirs of the American Entomological Institute 69. 824 pp. recorded H. zea from a species of Physalis, thus morphological characters should be carefully checked to separate larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
found on Physalis. Peterson (1962: L36) illustrated differences in the spinespine:
an unarticulated thornlike projection arising from the cuticle; compare to seta or spur
pattern of A4 which can be used to separate C. subflexa from H. zea. In addition, the SD2 setaseta:
a hairlike projection of the body wall that is articulated in a socket; compare to spine
is surrounded by a sclerotizedsclerotized:
hardened and tanned, so that it is yellow to black in color
area in C. subflexa but not in H. zea (Peterson 1962: L36, Wagner et al. 2011Wagner et al. 2011:
Wagner, D. L., D. F. Schweitzer, J. B. Sullivan and R. C. Reardon. 2011. Owlet caterpillars of eastern North America. Princeton University Press, New Jersey. 576 pp.).
The key by Hardwick (1965)Hardwick (1965):
Hardwick, D. F. 1965. The corn earworm complex. Memoirs of the Entomological Society of Canada 40: 1-247. is useful but not practical for APHIS. The first couplet lacks entries for Europe, Central America and the Caribbean. Granted, these areas can be treated with other existing keys. A bigger problem with his morphometric study is that the cuticle stretches between instars causing the pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
to shrink (Neunzig 1969:11). It is unknown if this affects the setal ratios that Hardwick (1965)Hardwick (1965):
Hardwick, D. F. 1965. The corn earworm complex. Memoirs of the Entomological Society of Canada 40: 1-247. used to recognize species, although he claimed a high success rate for the key.
Helicoverpa zea is highly polyphagous and widely distributed, but there are no records outside of the Americas, Hawaii and scattered collections on other Pacific Islands that may not have established (Hardwick 1965Hardwick 1965:
Hardwick, D. F. 1965. The corn earworm complex. Memoirs of the Entomological Society of Canada 40: 1-247.). The above diagnoses will allow species identifications from North America, Central America and the Caribbean.
It should be noted that H. phloxiphaga occurs only as far south as Mexico, and is quite rare in United States port interceptions. The other species (H. zea, C. virescens, and C. subflexa) are commonly intercepted.
Only the mid to last instars of H. zea can be identified by the mandible and cuticle microspinesmicrospines:
minute spines on the body, usually visible only under magnification
. First and second instars should be left at subfamily or consult Neunzig (1969)Neunzig (1969):
Neunzig, H. H. 1969. The biology of the tobacco budworm and the corn earworm in North Carolina with particular reference to tobacco as a host. North Carolina Agricultural Experiment Station Technical Bulletin 196. 76 pp. if there is a reason to make an identification of early instars of H. zea and C. virescens. For the quarantine decisions in the United States, both have the same action status, and this effort is usually not justified.
Identification of H. zea from South America is often not possible. One problem is the introduction of H. armigera to Brazil (Czepak et al. 2013Czepak et al. 2013:
Czepak, C., K. C. Albernaz, L. M. Vivan, H. O. Guimaraes and T. Carvalhais. 2013. Research note. First reported occurrence of Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae) in Brazil. Pesquisa Agropecuaria Tropical, Goiania 43: 110-113.), Paraguay, and Argentina (Murua et al. 2014Murua et al. 2014:
Murua, M. G., F. S. Scalora, F. R. Navarro, L. E. Cazado, A. Casmuz, M. E. Villagran, E. Lobos and G. Gastaminza. 2014. First record of Helicoverpa armigera (Lepidoptera: Noctuidae) in Argentina. Florida Entomologist 97: 854-856.). No morphological characters are yet known to separate the larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of H. armigera from H. zea; thus we can only go to genus Helicoverpa from these locations in South America.
Currently (June, 2014), it is possible to identify H. zea from Ecuador, Colombia, and Venezuela. There are sibling species of the zea complex in Peru and other parts of South America, especially H. gelotopoeon. For Peru and the rest of South America, it is best to say "Helicoverpa sp." Consult the Heliothinae guide (below) for details.
For Hawaii, if the interception is from corn, and the larvalarva:
the stages between the egg and pupa of those insects having complete metamorphosis
has conical pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
on A1, A2 and A8, then it can be called H. zea (Beardsley 1982Beardsley 1982:
Beardsley, J. W. Jr. 1982. A key to the late instar larvae of some Hawaiian Noctuidae. Proceedings of the Hawaiian Entomological Society 24: 37-49.). Helicoverpa hawaiiensis (Beardsley 1982Beardsley 1982:
Beardsley, J. W. Jr. 1982. A key to the late instar larvae of some Hawaiian Noctuidae. Proceedings of the Hawaiian Entomological Society 24: 37-49.) and some H. zea (SPIC) have flat pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
. Chloridea virescens also occurs in Hawaii (Beardsley 1982Beardsley 1982:
Beardsley, J. W. Jr. 1982. A key to the late instar larvae of some Hawaiian Noctuidae. Proceedings of the Hawaiian Entomological Society 24: 37-49.); it may be identified by the characters given above.
NOTE: Due to the continuing spread of H. armigera in South America, exercise caution when attempting identifications from that continent. There are no morphological characters to separate the larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of H. armigera from H. zea. When in doubt, default to "Helicoverpa sp." instead of attempting a species-level ID. The following keys may not reflect the most recent H. armigera distribution in South America.
Identification guide to larval Heliothinae (Lepidoptera: Noctuidae) of quarantine significance
Helicoverpa zea has been intercepted from the following locations:
Argentina, Barbados, Belize, Bolivia, Brazil, British Virgin Islands, Canada, Chile, Colombia, Costa Rica, Cuba, Dominica, Dominican Republic, Ecuador, El Salvador, Guatemala, Guyana, Haiti, Honduras, Jamaica, Mexico, Nicaragua, Panama, Peru, Puerto Rico, St. Lucia, Trinidad and Tobago, US Virgin Islands, Venezuela
Locations from outside of the Americas or the Caribbean have been omitted as these likely represent misidentifications.
Helicoverpa zea has been intercepted on the following hosts:
Abelmoschus esculentus, Abelmoschus sp., Allium fistulosum, Allium porrum, Allium sp., Aloe vera, Alstroemeria sp., Amaranthus caudatus, Amaranthus sp., Ananas comosus, Anethum graveolens, Annona cherimola, Antirrhinum majus, Antirrhinum sp., Apiaceae, Apium graveolens, Apium graveolens var. dulce, Apium sp., Artemisia dracunculus, Artocarpus altilis, Aster sp., Basilicum sp., Beta vulgaris var. cicla, Brassica chinensis, Brassica oleracea, Brassica oleracea var. botrytis, Brassica oleracea var. capitata, Brassica pekinensis, Brassica rapa, Brassica rapa ssp. chinensis, Brassica rapa ssp. pekinensis, Brassica sp., Cajanus cajan, Calendula officinalis, Calendula sp., Campanula sp., Capsicum annuum, Capsicum frutescens, Capsicum sinense, Capsicum sp., Carthamus sp., Chamaedorea sp., Chamomilla sp., Chenopodium album, Chenopodium berlandieri ssp nuttalliae, Chenopodium sp., Chichorium sp., Chicorum sp., Chrysanthemum sp., Cicer arietinum, Cicer sp., Cichorium intybus, Cichorium sp., Citrullus lanatus, Clematis sp., Colocasia sp., Corchorus capsularis, Coriandrum sativum, Cucumis sativus, Cucurbita maxima, Cucurbita pepo, Cucurbita sp., Cyamopsis tetragonoloba, Cynara cardunculus, Cynara scolymus, Daucus carota, Delphinium elatum, Delphinium sp., Dianthus caryophyllus, Dianthus sp., Equisetum sp., Eucalyptus sp., Fabaceae, Fragaria sp., Gerbera sp., Gladiolus sp., Gypsophila paniculata, Gypsophila sp., Helianthus annuus, Helianthus sp., Juglans sp., Lablab sp., Lactuca sativa, Lactuca sativa var. capitata, Lactuca sp., Lagenaria siceraria, Lagenaria sp., Leucadendron sp., Leucospermum sp., Limonium perezii, Limonium sp., Lippia sp., Luffa acutangula, Lycopersicon lycopersicum, Malus domestica, Mangifera indica, Matricaria recutita, Matricaria sp., Matthiola incana, Medicago sativa, Mentha arvensis, Mentha longifolia, Mentha piperita, Mentha sp., Mimosoideae, Moluccella sp., Momordica charantia, Momordica sp., Ocimum basilicum, Ocimum sp., Opuntia sp., Origanum majorana, Origanum sp., Origanum vulgare, Phaseolus sp., Phaseolus vulgaris, Pimenta sp., Pisum sativum, Pisum sativum var. macrocarpon, Pisum sp., Pithecellobium dulce, Poaceae, Polianthes tuberosa, Porophyllum sp., Portulaca oleracea, Pyrus communis, Raphanus sativus, Rosmarinus officinalis, Rubus sp., Saccharum officinarum, Salvia officinalis, Salvia sclarea, Salvia sp., Scabiosa sp., Solanaceae, Solanum integrifolium, Solanum lycopersicum var lycopersicum, Solanum melongena, Solanum sp., Solidago sp., Spinacia sp., Spiraea sp., Spondias purpurea, Suaeda sp., Tagetes erecta, Tagetes sp., Thymus sp., Thymus vulgaris, Vicia faba, Vigna sp., Yucca elephantipes, Zea mays, Zea sp., Zingiber sp.
There are several records in PestID of H. zea from Mexico on Physalis (Physalis ixocarpa, Physalis philadelphica, Physalis pubescens), some of these possibly represent interceptions of C. subflexa.
Click here to download a full-size printable PDF of this larval setal map
Click here to download a PDF of this fact sheet content and images