Subfamily: Mimosoideae.
Phylogenetic Number: 2.5.14.
Tribe: Ingeae.
Species Studied - Species in Genus: 1 studied; 1 in genus.
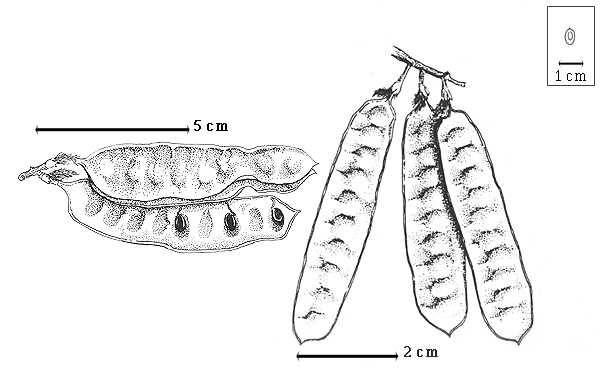
Fruit: A legume; unilocular; 7–12.5 cm long; 1.5–2.5 cm wide; 0.3–0.4 cm thick; 2–9 times longer than wide; with deciduous androecial sheath; with deciduous corolla; with deciduous calyx; without orifice formed by curving of fruit or fruit segments; straight; not plicate; not twisted; symmetrical; oblong to linear (broadly); not inflated; compressed; without beak; short tapered at apex to rounded at apex; aligned with longitudinal axis of fruit; short tapered at base; aligned with longitudinal axis of fruit; with the apex and base uniform in texture; coriaceous; seed chambers externally visible; margin not constricted to constricted; margin constricted along both margins (irregular once constricted); margin without sulcus; margin plain; wing(s) absent; substipitate; with all layers dehiscing (or tardily dehiscent), or indehiscent; splitting along suture(s). Dehiscence of valves along both sutures (splitting, remaining attached to sutures); apical and down; passive. Replum invisible. Epicarp dull; monochrome; brown; with surface texture uniform; glabrous to pubescent and indurate; with hairs erect; with simple hairs; eglandular; without spines; not smooth; with elevated features; veined, or not veined; transversely veined relative to fruit length (parallel reticulation tending to anastomose near midvalve); not tuberculate; not exfoliating; without cracks; without embedded tissue, much thicker than epicarp, running from base to apex. Mesocarp absent. Endocarp present; visible; dull; opaque; monochrome; tan; smooth; without adhering pieces of testa; septate to nonseptate; not exfoliating; entire. Seed(s) 1–14; length transverse to fruit length; neither overlapping nor touching; in 1 series. Funiculus up to 8 mm long; of 3 different lengths; thick; hooked to plicate. Aril absent.
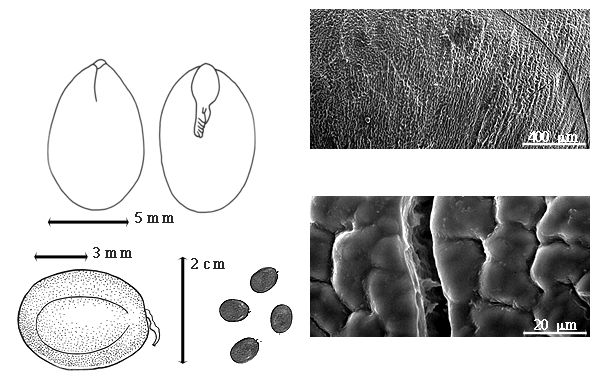
Seed: 4–13 mm long; 3–9 mm wide; 3–4 mm thick; not overgrown; not angular; symmetrical; ovate to elliptic, or oblong; compressed; with surface smooth; without visible radicle and cotyledon lobes; without hilar sinus; without umbo on seed faces; without medial ridge on each face. Cuticle not exfoliating; not inflated; not wrinkled. Testa present; without pieces of adhering epicarp; not adhering to endocarp; free from endocarp; glossy; not modified by a bloom; colored; monochrome; black; glabrous; not smooth; with recessed features; minutely pitted with small separate pits; osseous. Pleurogram present; 90 %. Pseudopleurogram absent. Fracture lines present. Rim absent. Wing(s) absent. Raphe not visible. Hilum present; fully concealed; concealed by funicular remnant; without faboid split; punctiform; apical at apex of radicle tip; flush; not within corona, halo, or rim. Lens discernible; 0.3–0.4 mm long; with margins curved; elliptic to circular; not in groove of raphe; mounded; same color as testa; black; not within corona, halo, or rim. Endosperm present; thick; not pluglike and not resembling tip of radicle; adnate to testa. Cotyledons smooth; both outer faces flat; both the same thickness; both more or less of equal length; not folded; margin entire 180 degrees from base of radicle; similar at apex; partially concealing radicle; split over radicle; with the interface division terminating at base of radicle; without margins recessed; inner face flat; glabrous on inner face. Embryonic axis straight; parallel to length of seed; without a joint evident between the radicle and the cotyledons. Radicle differentiated from cotyledon; straight with embryonic axis; centered between cotyledons. Plumule well developed; glabrous.
Dell (1980) demonstrated that entry of water into heated hard seeds of P. lophantha (Albizia lophantha (C.L. von Willdenow) G. Bentham of Dell) is controlled by eruption of a small (0.3 X 0.25 mm) strophiolar plug adjacent to the hilum. This plug is the lens. Nielsen et al. (1983a, 1983b) revised the genus. Barneby and Grimes (1996) elevated Nielsen's Paraserianthes sect. Falcataria I.C. Nielsen to generic rank thereby making Paraserianthes monotypic. Their desciption of the genus covers only P. lophantha var. lophantha which is native to southwestern Australia.
Authors: J.H. Kirkbride, Jr., C.R. Gunn, A.L. Weitzman, M.J. Dallwitz, K.R. Thiele
Content last updated April 2003
Released April 2024 on idtools.org