
probably harmful; mites presumably kill bee larvae and eggs in infested cells
Horstia Oudemans, 1900
Superorder Acariformes » Order Sarcoptiformes » Suborder Oribatida » Infraorder Desmonomata » Hyporder Astigmata » Family Acaridae » Genus Horstia
Trichotarsus ornatus Oudemans, 1900
Ceroglyphus Vitzthum, 1920Vitzthum, 1920:
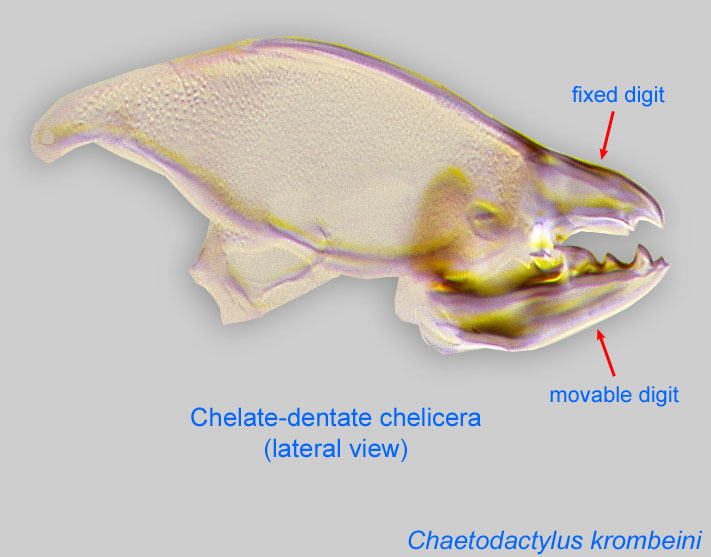
Vitzthum, H. G. 1920[1918]. Acarologische Beobachtungen. Zweite Reihe. Archiv für Naturgeschichte . 84A : 1-40. [1918] (described from males with enlarged cheliceraechelicera:
Anterior, paired appendage of the body. Primary organ for food acquisition, adapted for chewing, piercing, tearing, sucking, or filtering.
 ); Petzschia Oudemans, 1923
); Petzschia Oudemans, 1923
Phoretic deutonymphphoretic deutonymph:
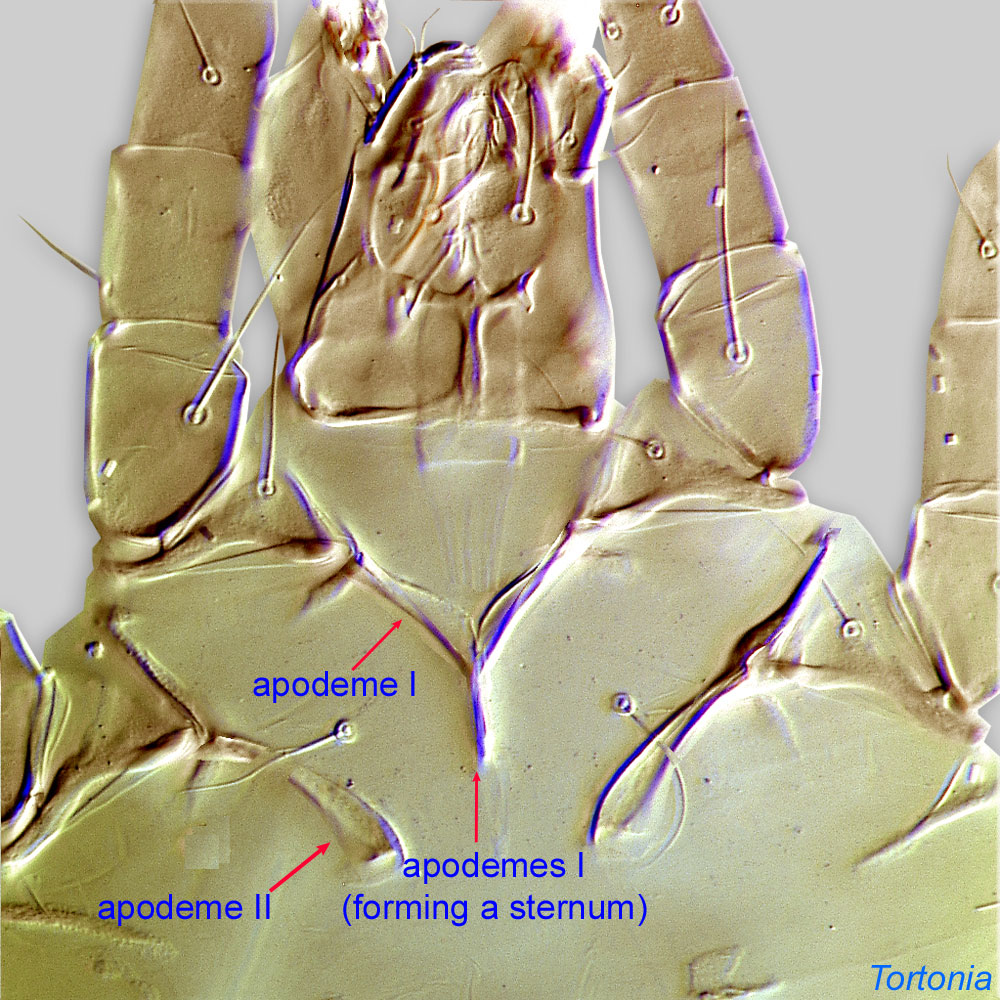
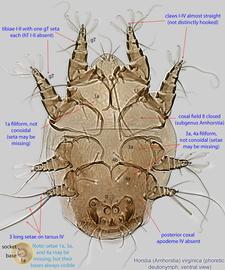
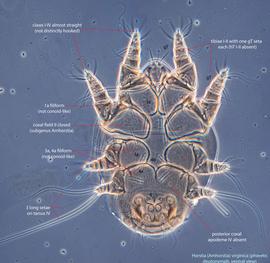
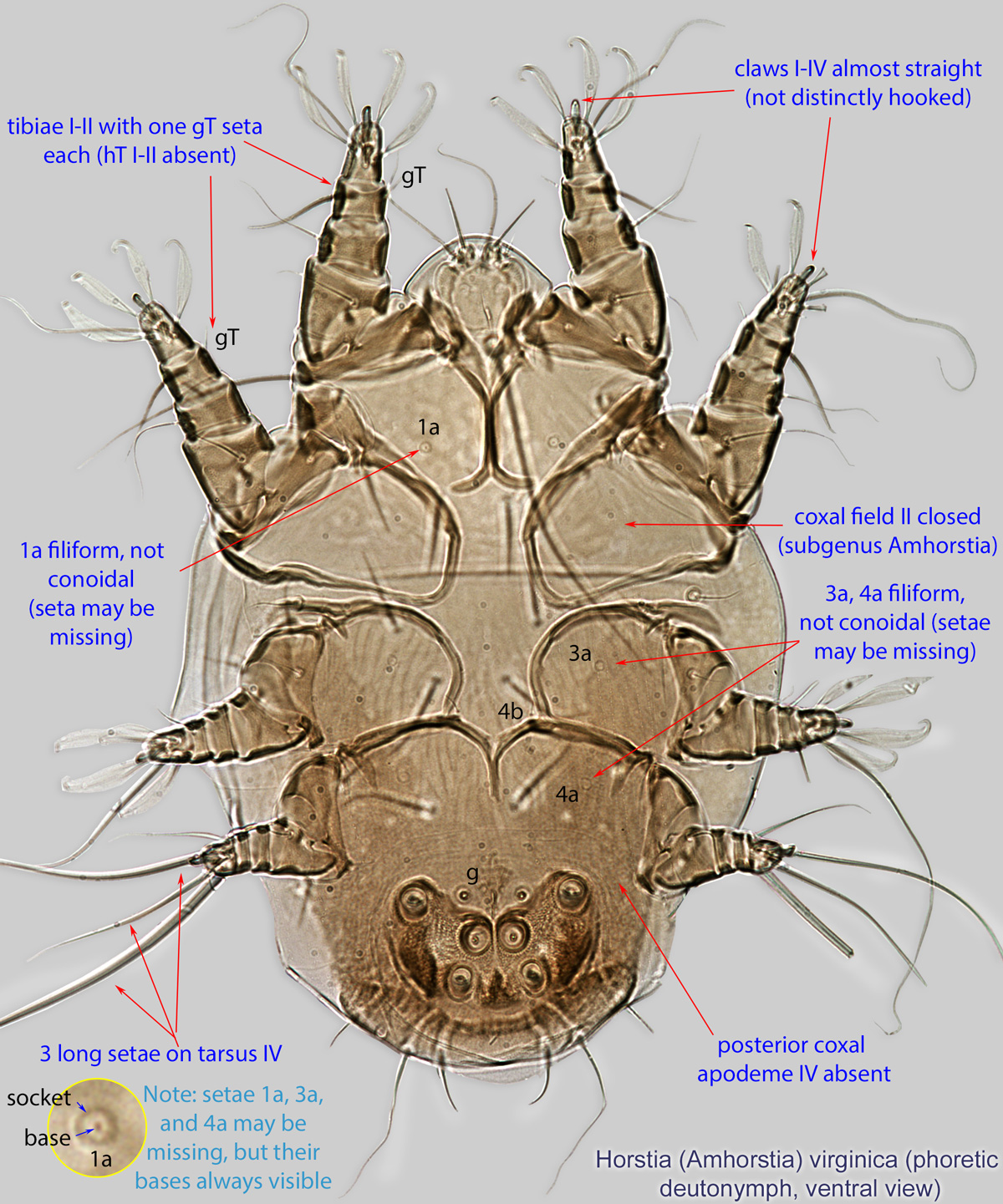
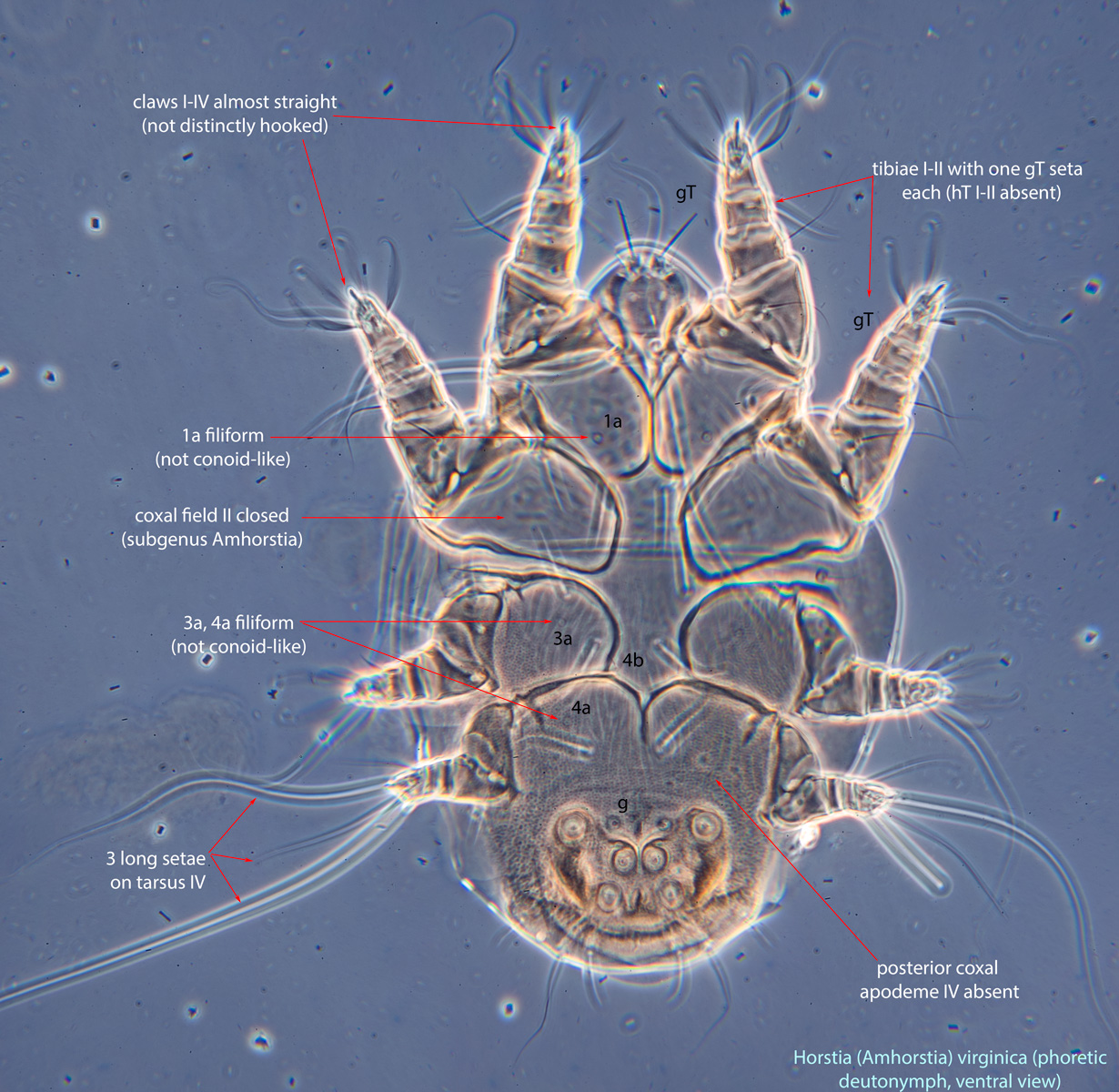
An active nymphal stage in Astigmata that serves for dispersal on arthropod or vertebrate hosts. A subtype of heteromorphic deutonymph. See Life stages page for more details. : Coxal setae 1a, 3a, 4a filiform (not conoidal) (Figs. 3, 4). Posterior coxal apodemeapodeme:
: Coxal setae 1a, 3a, 4a filiform (not conoidal) (Figs. 3, 4). Posterior coxal apodemeapodeme:
Internal sclerite that serves as an attachment site for muscles. Most commonly used (as "coxal apodeme") to describe elements of coxae fused to the ventral body in Acariformes (coxae are free and not fused to the body in Parasitiformes), and may be variously referred to as ventral, sternal, anterior, or posterior.
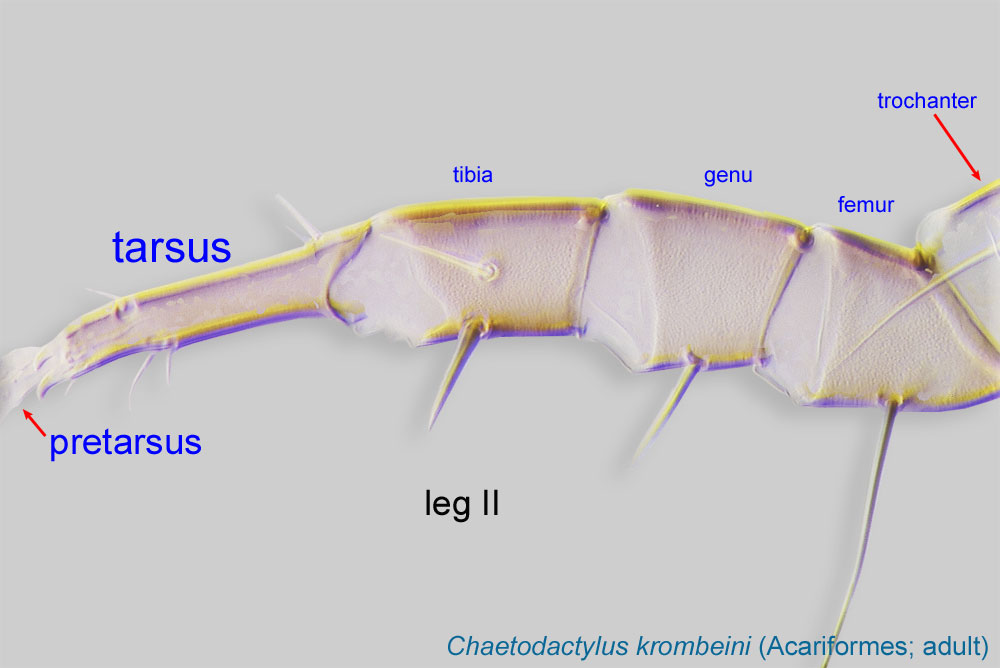
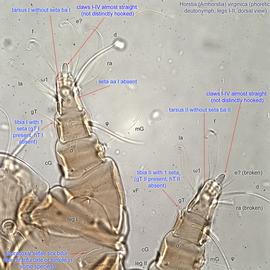
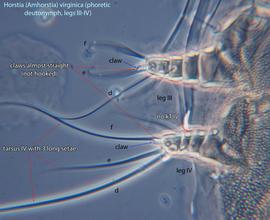
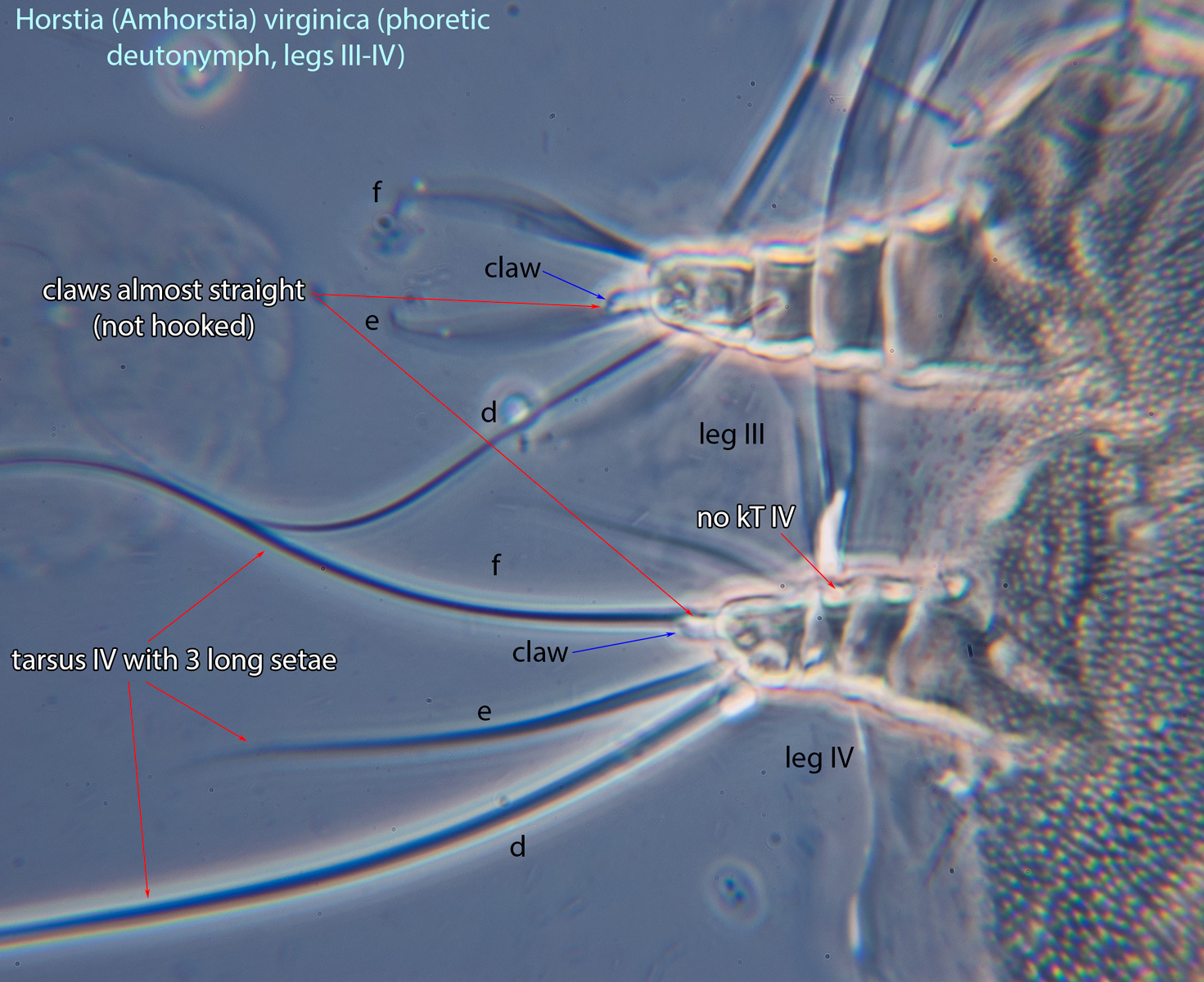
 IV absent (Figs. 3, 4). Supracoxal setae scx may be bifurcate (Figs. 5,11) or trifurcate. Claws I-IV almost straight (not distinctly hooked) (Figs. 3, 4, 5, 6). Tarsitarsus:
IV absent (Figs. 3, 4). Supracoxal setae scx may be bifurcate (Figs. 5,11) or trifurcate. Claws I-IV almost straight (not distinctly hooked) (Figs. 3, 4, 5, 6). Tarsitarsus:
Terminal segment (also known as podomere or palpomere) of legs or palps. In Parasitoformes it can be subdivided into telotarsus and basitarsus.
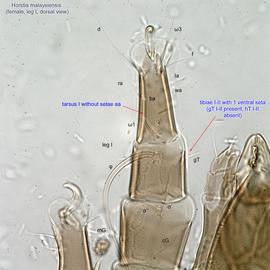
 I-II without setae ba I-II (Fig. 5). Tarsustarsus:
I-II without setae ba I-II (Fig. 5). Tarsustarsus:
Terminal segment (also known as podomere or palpomere) of legs or palps. In Parasitoformes it can be subdivided into telotarsus and basitarsus.
I without seta aa (Fig. 5). Tarsustarsus:
Terminal segment (also known as podomere or palpomere) of legs or palps. In Parasitoformes it can be subdivided into telotarsus and basitarsus.
IV with 3 long setae (Figs. 3, 4, 6). Tibiaetibia:
Leg or palp segment (also known as podomere or palpomere) between tarsus and genu.
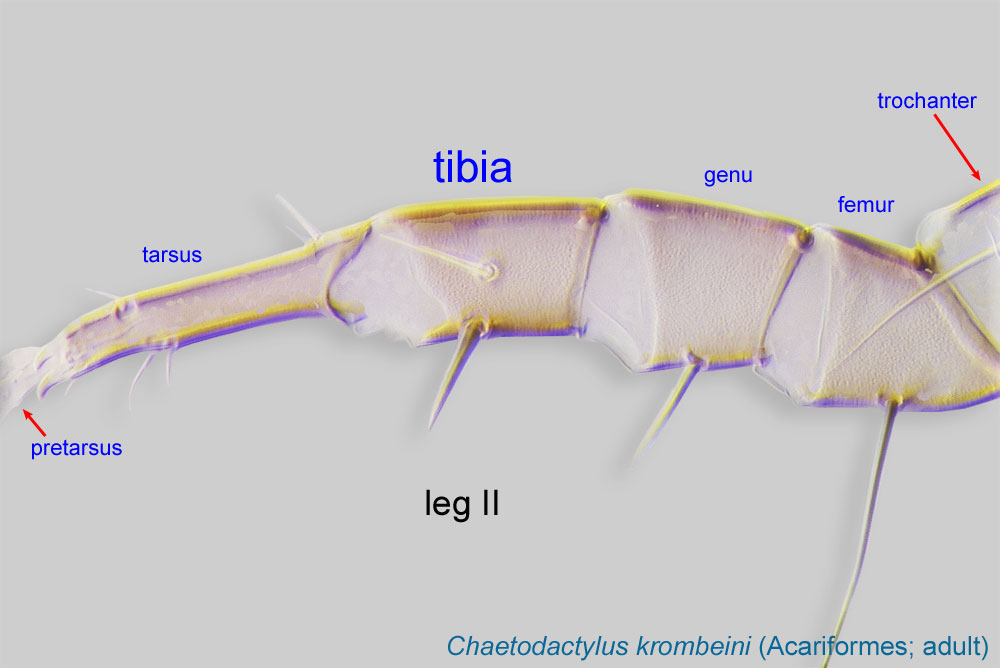 I-II with 1 seta each (gT I-II present, hT I-II absent) (Figs. 3, 4, 5). Tibiaetibia:
I-II with 1 seta each (gT I-II present, hT I-II absent) (Figs. 3, 4, 5). Tibiaetibia:
Leg or palp segment (also known as podomere or palpomere) between tarsus and genu.
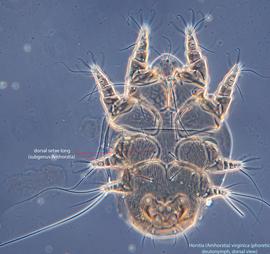
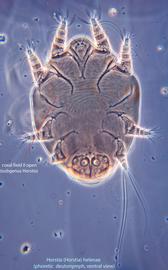
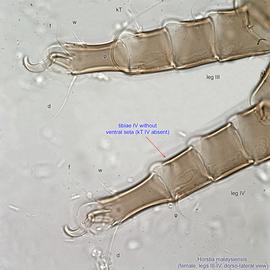
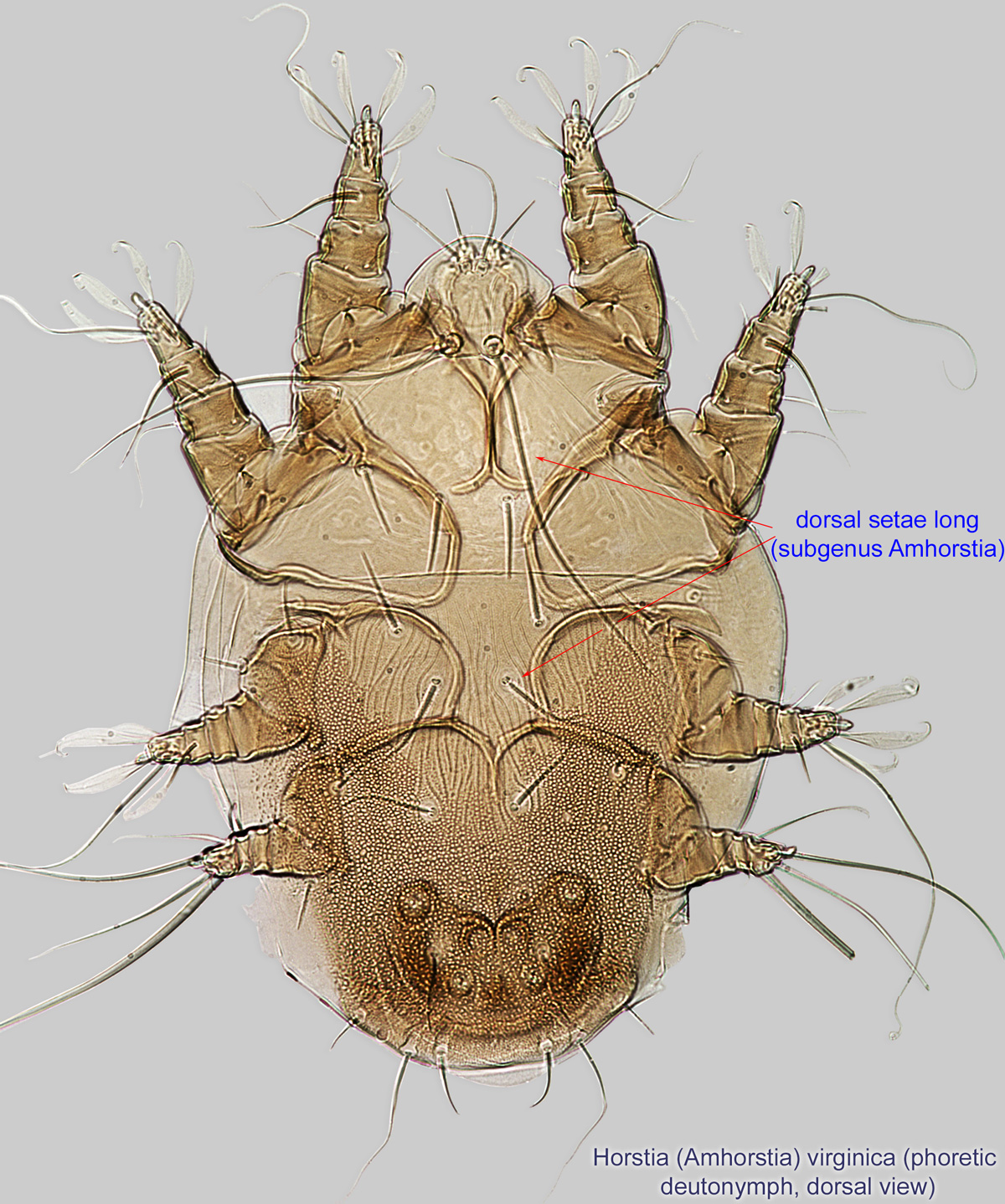
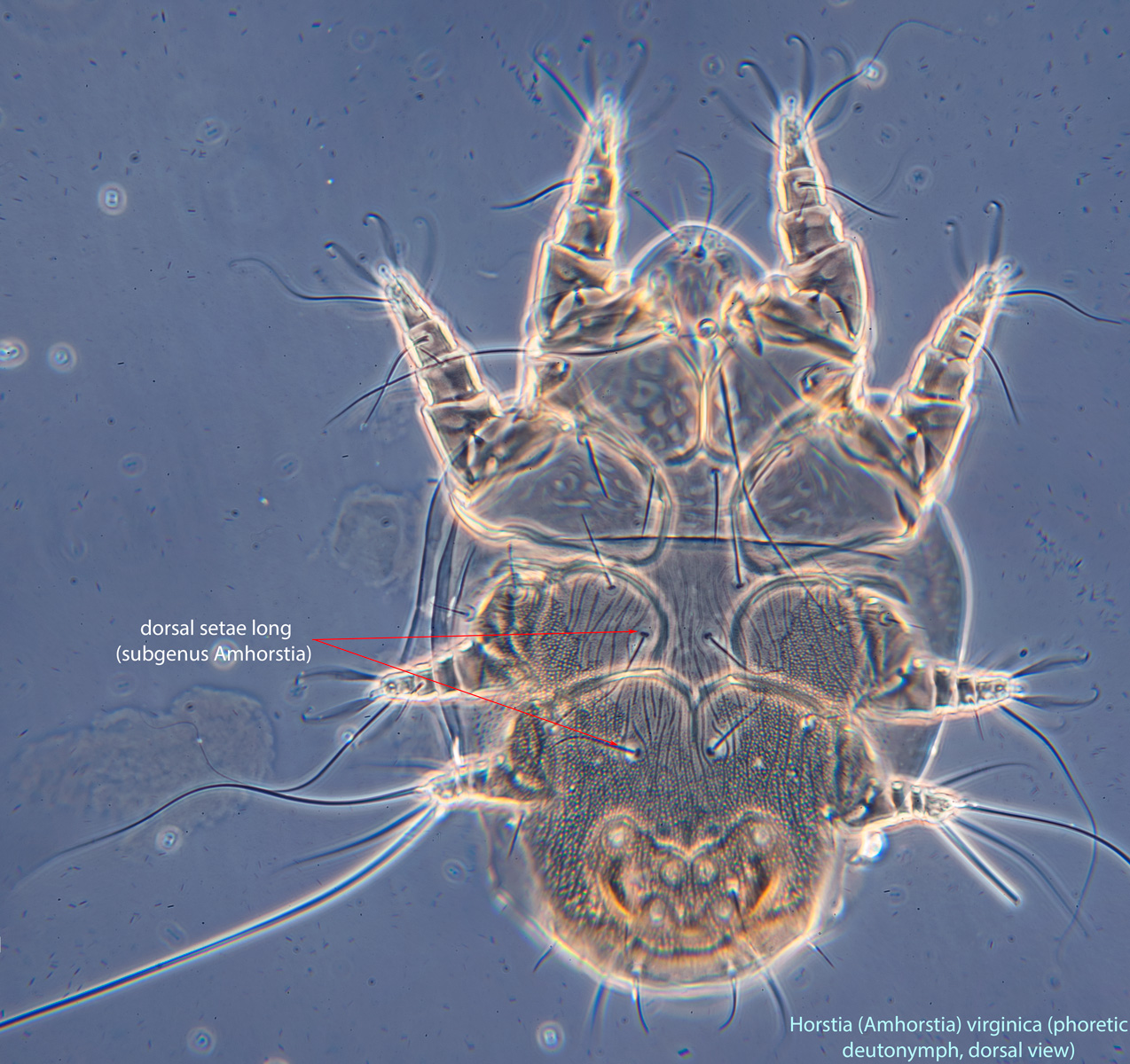
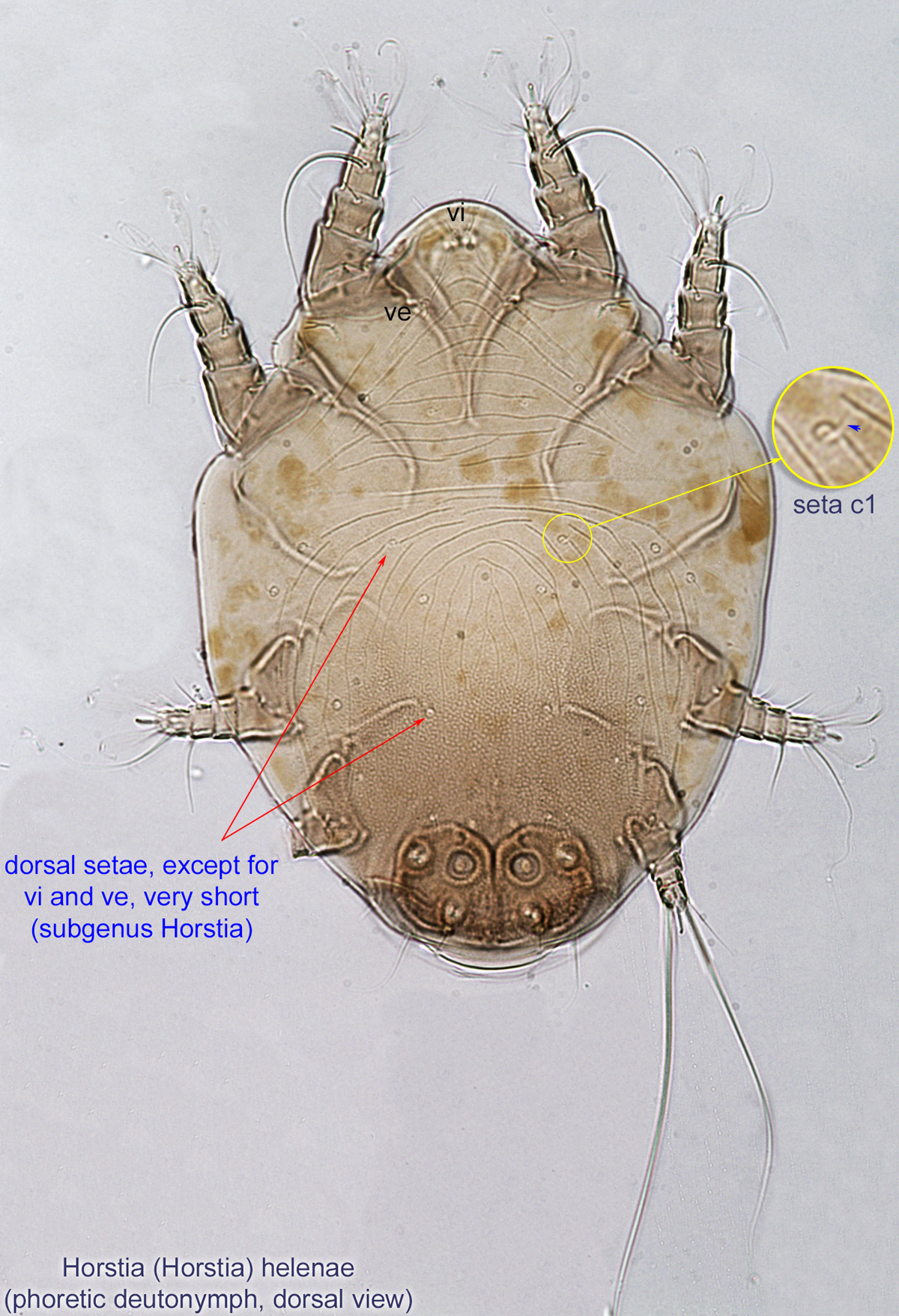
IV without ventral seta (kT IV absent) (Fig. 6). Dorsal setae long (subgenus Amhorstia, Figs. 1, 2) or very short (subgenus Horstia, Figs. 7, 8). Coxal fields II closed (subgenus Amhorstia, Figs. 3, 4) or open (subgenus Horstia, Figs. 9,10).
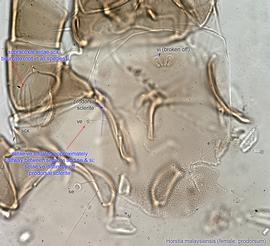
Adult: In female, setae ve situated posterior to setae vi, approximately halfway between vi and se & si, distinctly off prodorsalprodorsal:
Pertaining to the prodorsum.
scleritesclerite:
A component section of an exoskeleton; a plate forming the skeleton of an arthropod.
(Fig. 15). Supracoxal setae scx may be bifurcate (Fig. 15). Tarsustarsus:
Terminal segment (also known as podomere or palpomere) of legs or palps. In Parasitoformes it can be subdivided into telotarsus and basitarsus.
I without setae aa (Fig. 16). Tibiaetibia:
Leg or palp segment (also known as podomere or palpomere) between tarsus and genu.
I-II with 1 ventral seta (gT I-II present, hT I-II absent) (Fig. 16). Tibiaetibia:
Leg or palp segment (also known as podomere or palpomere) between tarsus and genu.
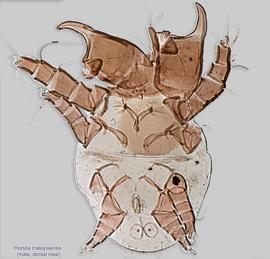
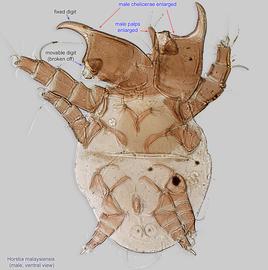
IV without ventral seta (kT IV absent) (Fig. 17). Male sometimes with enlarged cheliceraechelicera:
Anterior, paired appendage of the body. Primary organ for food acquisition, adapted for chewing, piercing, tearing, sucking, or filtering.
and palpspalp:
Second (after chelicera) paired appendage of the gnathosoma. Has a sensory function, but may be variously modified for other functions (e.g., raptorial, attachment to host, or filtering).
 (Figs. 13, 14).
(Figs. 13, 14).
A dichotomous key to phoretic deutonymphsphoretic deutonymph:
An active nymphal stage in Astigmata that serves for dispersal on arthropod or vertebrate hosts. A subtype of heteromorphic deutonymph. See Life stages page for more details. is available in Fain, 1984Fain, 1984:
Fain, A. 1984. Notes sur les hypopes du genre Horstia Oudemans, 1905 (Acari, Acaridae), phoretiques sur les hymenopteres. Acarologia . 25 : 259-270.. Two species, Horstia major and H. minor, were described later and should be identified using their original description (Alzuet and Abrahamovich, 1987Alzuet and Abrahamovich, 1987:
Alzuet, A. B. d. amp; A. H. Abrahamovich. 1987 [1985]. Deutoninfas (hypopi) de los géneros Sennertia Oudemans, 1905 y Horstia Oudemans, 1905 (Acari: Astigmata) sobre Xylocopa ( S .) splendidula splendidula Lepeletier, 1841 (Hymenoptera: Apoidea). Revista de la Sociedad Entomologica Argentina . 44 : 345-351.).
Adults are known for H. virginica, H. longa, H. monstruosa, and H. malaysiensis (probably a junior synonym of H. helenae), and these should be identified using the original descriptions.
SE Palaearctic (Japan), Nearctic, Neotropical, Afrotropical, Oriental, and Australian regions. The subgenus Amhorstia is distributed in The New World, while the subgenus Horstia is distributed in the Old World.
Associated with large carpenter bees (Xylocopa). Occasional or accidental records include small carpenter bees (Ceratina), bumble bees (Bombus), and leafcutting and resin bees (Megachile).
Permanentpermanent:
associated exclusively with bees or their close relative, wasps; cannot live without these hosts
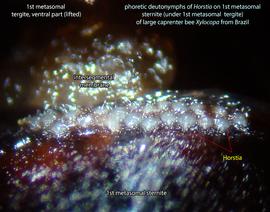
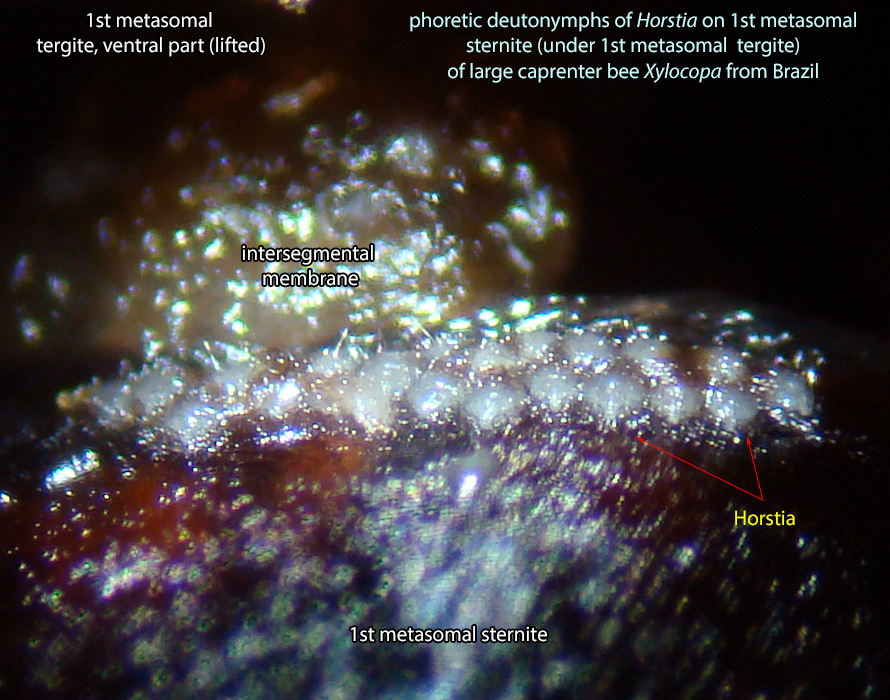
disperse on bee hosts. Mites are usually situated in intersegmental space, on the sides of first metasomal sternite (hidden by overlapping tergite 1) on bees from the Neotropical region (Figs. 18, 19).Biological observations on Horstia virginica associated with Xylocopa virginica krombeini are summarized below from Krombein, 1962bKrombein, 1962b:
Krombein, K. V. 1962b. Biological notes on acarid mites associated with solitary wood-nesting wasps and bees. Proceedings of the Entomological Society of Washington . 64 : 11-19.; Krombein, 1967Krombein, 1967:
Krombein, K. V. 1967. Trap-nesting wasps and bees: life histories, nests, and associates. Washington, D.C: Smithsonian press. 570 pp.:
All examined nest cells infested with Horstia contained dead bee larvae and some infested cells contained dead eggs. Presumably the mites caused the death of the bees, but the exact mechanism is unknown. Occasionally bees have been able to develop successfully in mite-infested cells, but these bee larvae may have reached maturity before their cells were invaded by mites. An alternate explanation for these observations is that mites may disperse relatively easily through a nesting aggregation, which may allow mites to exploit those cells where bees have not developed due to other mortality factors and still allow deutonymphsdeutonymph:
Ontogenetic stage between protonymph and tritonymph (or adult, if tritonymph is absent). See Life stages page for more details. to locate and attach to live emerging bees.
Rearing experiments demonstrated that mites fed on nectar (and possibly also the upper layer of pollen grains) and flourished on this food source without the presence of either live or dead bees.
Presumably bees from non-infested cells would become infested as they passed through an infested cell during their emergence from the nest. Or, it may be that live phoretic deutonymphsphoretic deutonymph:
An active nymphal stage in Astigmata that serves for dispersal on arthropod or vertebrate hosts. A subtype of heteromorphic deutonymph. See Life stages page for more details. remain in the old nest and infest new cells when another female uses the old nest.
Phoretic deutonymphsphoretic deutonymph:
An active nymphal stage in Astigmata that serves for dispersal on arthropod or vertebrate hosts. A subtype of heteromorphic deutonymph. See Life stages page for more details. have been found clustering on the female bee pupa on the pupa's thoracic sternum, middle of mesonotum, and wings.
Our observations indicate that phoretic deutonymphsphoretic deutonymph:
An active nymphal stage in Astigmata that serves for dispersal on arthropod or vertebrate hosts. A subtype of heteromorphic deutonymph. See Life stages page for more details. of Horstia are very common in intersegmantal spaces on the sides of first metasomal sternite (hidden by overlapping tergite 1) of large carpenter bees Xylocopa in the Neotropics (Figs. 18, 19).
Authors: P. Klimov, B. OConnor, R. Ochoa, G. Bauchan, A. Redford, J. Scher
Last updated October 2016
tool images at ITP Node
idtools.org









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}