Subfamily: Faboideae.
Phylogenetic Number: 3.7.
Tribe: Millettieae.
Species Studied - Species in Genus: 3 studied; 6 in genus.
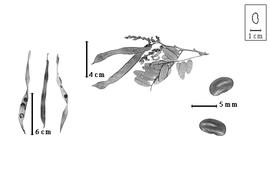
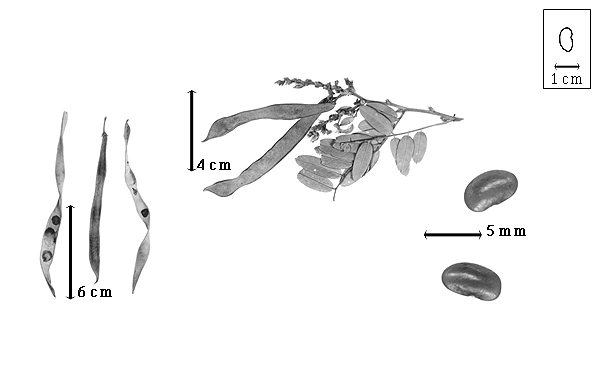
Fruit: A legumelegume:
usually dry, dehiscent fruit derived from a single carpel that opens along two longitudinal sutures.jpg) ; unilocular; 6–11 cm long; 0.5–0.9 cm wide; 0.2–0.3 cm thick; more than 9 times longer than wide; with deciduous androecial sheath; with deciduous corolla; with persistent calyx; with calyx shorter than fruit; without orifice formed by curving of fruit or fruit segments; straight; not plicate; not twisted; asymmetrical; falcate, or irregularly fusiform; with both sutures parallelly curved, or both sutures unequally curved; not inflated; compressed to flattened; without beak, or with beak; declined; with solid beak the same color and texture as fruit; long tapered at apex; oblique with longitudinal axis of fruit; long tapered at base to tapered at base; aligned with longitudinal axis of fruit, or oblique with longitudinal axis of fruit; with the apex and base uniform in texture; coriaceous; seed chambers externally visible, or invisible; margin constricted, or not constricted; margin slightly constricted along both margins; margin without sulcus; margin plain; nonstipitate; with all layers dehiscing; splitting along suture(s). Dehiscence of valves along both sutures; apical and down; active; with valves twisting. Replum invisible. Epicarp dull; monochrome; brown, or gray; with surface texture uniform; pubescent and indurate; with hairs erect; with 1 type of pubescence, or 2 types of pubescence; pilose to velutinous to villous; with pubescence brown, or tan, or white; with long white and short golden hairs intermixed, or contiguous areas of golden and white hairs; with pubescence uniformly distributed; with simple hairs; stiff; with hair bases plain; eglandular; without spines; smooth, or not smooth; with elevated features; not veined; not tuberculate; wrinkled; not exfoliating; without cracks; without embedded tissue, much thicker than epicarp, running from base to apex. Mesocarp present; thick; surface not veined; 2-layered; without balsamic vesicles; without fibers; without reniform canals; with vitriol layer over solid layer, or spongy layer over solid layer; coriaceous. Endocarp present; visible; dull; opaque; monochrome, or mottled; tan; with mottling over seed chambers; with brown overlay; smooth and floury-filamentous; without adhering pieces of testa; subseptate; with septa eglandular; chartaceous; exfoliating in part, or not exfoliating; remaining fused to mesocarp and epicarp; entire. Seed(s) 6–10; length parallel with fruit length, or oblique to fruit length; neither overlapping nor touching; in 1 series. Funiculus 1–2 mm long; of 1 length only; flattened; straight. Aril present; dry; when dry very small rim-aril; entire; covering less than 1/2 of seed; white.
; unilocular; 6–11 cm long; 0.5–0.9 cm wide; 0.2–0.3 cm thick; more than 9 times longer than wide; with deciduous androecial sheath; with deciduous corolla; with persistent calyx; with calyx shorter than fruit; without orifice formed by curving of fruit or fruit segments; straight; not plicate; not twisted; asymmetrical; falcate, or irregularly fusiform; with both sutures parallelly curved, or both sutures unequally curved; not inflated; compressed to flattened; without beak, or with beak; declined; with solid beak the same color and texture as fruit; long tapered at apex; oblique with longitudinal axis of fruit; long tapered at base to tapered at base; aligned with longitudinal axis of fruit, or oblique with longitudinal axis of fruit; with the apex and base uniform in texture; coriaceous; seed chambers externally visible, or invisible; margin constricted, or not constricted; margin slightly constricted along both margins; margin without sulcus; margin plain; nonstipitate; with all layers dehiscing; splitting along suture(s). Dehiscence of valves along both sutures; apical and down; active; with valves twisting. Replum invisible. Epicarp dull; monochrome; brown, or gray; with surface texture uniform; pubescent and indurate; with hairs erect; with 1 type of pubescence, or 2 types of pubescence; pilose to velutinous to villous; with pubescence brown, or tan, or white; with long white and short golden hairs intermixed, or contiguous areas of golden and white hairs; with pubescence uniformly distributed; with simple hairs; stiff; with hair bases plain; eglandular; without spines; smooth, or not smooth; with elevated features; not veined; not tuberculate; wrinkled; not exfoliating; without cracks; without embedded tissue, much thicker than epicarp, running from base to apex. Mesocarp present; thick; surface not veined; 2-layered; without balsamic vesicles; without fibers; without reniform canals; with vitriol layer over solid layer, or spongy layer over solid layer; coriaceous. Endocarp present; visible; dull; opaque; monochrome, or mottled; tan; with mottling over seed chambers; with brown overlay; smooth and floury-filamentous; without adhering pieces of testa; subseptate; with septa eglandular; chartaceous; exfoliating in part, or not exfoliating; remaining fused to mesocarp and epicarp; entire. Seed(s) 6–10; length parallel with fruit length, or oblique to fruit length; neither overlapping nor touching; in 1 series. Funiculus 1–2 mm long; of 1 length only; flattened; straight. Aril present; dry; when dry very small rim-aril; entire; covering less than 1/2 of seed; white.
Seed: 3.5–6 mm long; 2–4 mm wide; 2–2.5 mm thick; not overgrown; not angular; symmetrical; reniform; compressed to terete (nearly); with surface smooth; without visible radicle and cotyledon lobes; with shallow hilar sinus; without umbo on seed faces; without medial ridge on each face. Testa present; without pieces of adhering epicarp; not adhering to endocarp; free from endocarp; dull; not modified by a bloom; colored; monochrome, or mottled; with infrequent mottles; brown; with brown overlay (darker); glabrous; smooth; chartaceous. Pleurogram absent. Pseudopleurogram absent. Fracture lines absent. Rim absent. Wing(s) absent. Raphe not visible. Hilum present; visible; with faboid split; with the lips of the faboid split lighter colored than the rest of the hilum and therefore conspicuous; larger than punctiform; 0.5–0.8 mm long; with curved outline; oval; apical according to radicle tip but marginal according to seed length; recessed; within halo; halo darker than testa. Lens discernible; ca. 1 mm long; with margins straight, or curved; linear; hourglass or dumbbell-shaped; not in groove of raphe; confluent with hilum; slightly mounded; similar color as testa; darker than testa; brown; not within corona, halo, or rim. Endosperm absent. Cotyledons smooth; both outer faces convex; both the same thickness; both more or less of equal length; not folded; margin entire 180 degrees from base of radicle; similar at apex; partially concealing radicle; notched at radicle; without lobes; with the interface division terminating at base of radicle; without margins recessed; white; inner face flat, or wrinkled; glabrous on inner face. Embryonic axis oblique; parallel to length of seed; without a joint evident between the radicle and the cotyledons. Radicle differentiated from cotyledon; bulbose; lobe tip curved; oblique to cotyledons; centered between cotyledons; less than 1/2 length of cotyledons. Plumule rudimentary; glabrous.
Madagascar.
Old World; Madagascar.
This genus was described and monographed by Du Puy and Labat (1995). They (1995) stated, "this genus appears to be somewhat intermediate between Mundulea and Chadsia, with P. ambatoana forming the closest link to the latter.".
Tribe Millettieae
Geesink (1981) treated this tribe, as others before him, under the tribal name Tephroseae, but we now know that its correct name is Millettieae. Recent tribal studies (Geesink, 1981, 1984; Polhill, 1994a, 1994b) have arranged the genera in alphabetical order without phylogenetic numbers. Geesink (1984) monographed tribe Millettieae, and presented descriptive notes about fruits and seeds and in situ fruit and seed drawings. However, we are not entirely following Geesink (1984) for generic parameters because he either questioned the status of many of his new genera or did not make the necessary species transfers. The few new genera which he clearly recognized are being accepted. Lavin et al. (1998) developed a preliminary infratribal classification of six informal groups using phytochrome nucleotides: Millettia group, Lonchocarpus group, Derris group, Tephrosia group, "primitive" group, and Phaseoleae group. Lavin (1987) transferred Sphinctospermum to Millettieae. Lavin and Doyle (1991) carried out cladistic analyses integrating morphological and chloroplast DNA data, and concluded that it is a member of Robineae where we have placed it (now 8.12).
Authors: J.H. Kirkbride, Jr., C.R. Gunn, A.L. Weitzman, M.J. Dallwitz, K.R. Thiele
Content last updated April 2003
Released April 2024 on idtools.org


{kind=link}
{kind=link}