Subfamily: Faboideae.
Phylogenetic Number: 3.7.
Tribe: Millettieae.
Species Studied - Species in Genus: 3 studied; 12 in genus.
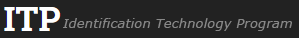
Fruit: A legume; unilocular; (4.5–)6–12 cm long; 1.5–3 cm wide; 0.2–0.6 cm thick; 2–9 times longer than wide; with deciduous androecial sheath; with deciduous corolla; with persistent calyx; with calyx shorter than fruit; without orifice formed by curving of fruit or fruit segments; straight to curved; not plicate; not twisted; symmetrical; oblong to elliptic (when fruit short); not inflated; flattened; with beak; straight; with solid beak the same color and texture as fruit; long tapered at apex to tapered at apex to short tapered at apex; aligned with longitudinal axis of fruit to oblique with longitudinal axis of fruit; long tapered at base to tapered at base, or rounded at base; aligned with longitudinal axis of fruit; with the apex and base uniform in texture; membranous to chartaceous; seed chambers externally visible; seed chambers with the raised seed chambers not torulose; margin not constricted, or constricted (rarely, where seeds aborted); margin constricted along both margins; margin without sulcus; margin embellished, or plain; margin with thickened sutural areas, or wing(s) (just one species winged, Geesink, 1984Geesink, 1984:
Geesink R. 1984. Scala Millettiearum. E.J. Brill/Leiden University Press, Leiden, The Netherlands.); wing(s) absent, or present (just one species winged, Geesink, 1984); wing(s) 1; wing(s) sutural; wing(s) on 1 suture (on upper suture, Geesink, 1984); stipitate; with the stipe 4–7 mm long; indehiscent. Replum invisible. Epicarp dull; monochrome; brown, or tan, or gray (greenish); with surface texture uniform; glabrous; eglandular; without spines; not smooth; with elevated features; veined; reticulately veined; not tuberculate; not exfoliating; without cracks; without embedded tissue, much thicker than epicarp, running from base to apex. Mesocarp absent. Endocarp present; visible; dull; opaque; monochrome; tan; smooth; without adhering pieces of testa; nonseptate; chartaceous; not exfoliating; remaining fused to epicarp; without wings; entire. Seed(s) 1–4(–8); length parallel with fruit length; neither overlapping nor touching; in 1 series. Funiculus 0.3 mm long; of 1 length only; filiform; straight. Aril present, or absent; dry; when dry rim-aril; entire; covering less than 1/2 of seed; without tongue (or flap) on lips of 2-lipped rim-aril; cream.
Seed: 9–15 mm long; 6–10 mm wide; 2–4 mm thick; not overgrown; not angular; symmetrical; reniform to oblong; compressed to flattened; with surface wrinkled; without visible radicle and cotyledon lobes; with deep hilar sinus; without umbo on seed faces; without medial ridge on each face. Cuticle not exfoliating; not inflated; not wrinkled. Testa present; with pieces of adhering epicarp; not adhering to endocarp; free from endocarp; glossy to dull; not modified by a bloom; colored; monochrome; reddish to dark brown; glabrous; smooth; chartaceous. Pleurogram absent. Pseudopleurogram absent. Fracture lines absent. Rim absent. Wing(s) absent. Raphe visible, or not visible (when testa dark brown); from hilum through lens and terminating before base of seed; not bifurcating; color of testa, or darker than testa; dark brown; flush. Hilum present; visible; with faboid split; with the lips of the faboid split the same color as the rest of the hilum; larger than punctiform; 0.8–1 mm long; with curved outline; elliptic; apical according to radicle tip but marginal according to seed length; recessed; not within corona, halo, or rim. Lens discernible; 0.8–1 mm long; with margins straight; triangular; not in groove of raphe; adjacent to hilum; ca. 0.2 mm from hilum; flush; similar color as testa; darker than testa; reddish to dark brown; not within corona, halo, or rim. Endosperm present; thin; not pluglike and not resembling tip of radicle; covering entire embryo, or covering at least 1/2 of embryo, but not entire embryo; adnate to testa. Cotyledons smooth; both outer faces convex; both the same thickness; both more or less of equal length; not folded; margin entire 180 degrees from base of radicle; similar at apex; not concealing radicle; entire over radicle; without lobes; with the interface division terminating at base of radicle; without margins recessed; tan; inner face flat; glabrous on inner face. Embryonic axis oblique; oblique to length of seed; without a joint evident between the radicle and the cotyledons. Radicle differentiated from cotyledon; bulbose; lobe tip straight; oblique to cotyledons; centered between cotyledons; less than 1/2 length of cotyledons. Plumule moderately developed; glabrous.
Africa and Madagascar.
Old World; Africa and Madagascar (Schrire, 2000).
Traditionally this genus has been included in Lonchocarpus (3.7). Geesink (1984) accepted it with 15 African and four American species, but the botanical community continued to include it in Lonchocarpus. Schrire (2000a, 2000b) also accepted the genus and treated its African and Madagascarian species, so we are now following Schrire. He maintained the four American species in Lonchocarpus, so this genus is strictly African and Madagascarian. Du Puy et al. (2002) did not mention or cite Philenoptera in The Leguminosae of Madagascar and accepted Lonchocarpus madagascariensis (Vatke) Polhill, which Schrire (2000) treated as P. madagascariensis (Vatke) Schrire.
Tribe Millettieae
Geesink (1981) treated this tribe, as others before him, under the tribal name Tephroseae, but we now know that its correct name is Millettieae. Recent tribal studies (Geesink, 1981, 1984; Polhill, 1994a, 1994b) have arranged the genera in alphabetical order without phylogenetic numbers. Geesink (1984) monographed tribe Millettieae, and presented descriptive notes about fruits and seeds and in situ fruit and seed drawings. However, we are not entirely following Geesink (1984) for generic parameters because he either questioned the status of many of his new genera or did not make the necessary species transfers. The few new genera which he clearly recognized are being accepted. Lavin et al. (1998) developed a preliminary infratribal classification of six informal groups using phytochrome nucleotides: Millettia group, Lonchocarpus group, Derris group, Tephrosia group, "primitive" group, and Phaseoleae group. Lavin (1987) transferred Sphinctospermum to Millettieae. Lavin and Doyle (1991) carried out cladistic analyses integrating morphological and chloroplast DNA data, and concluded that it is a member of Robineae where we have placed it (now 8.12).
|
Fruit and seed: P. cyanescens (J.H.K. Schumann & P. Thonning) G.E. Roberty - bottom fruit in left fruit grouping; P. laxiflora (J.P.A. Guillemin) G.E. Roberty - middle fruit in left fruit grouping; P. violacea (J.F. Klotzsch) B.D. Schrire - upper fruit in left fruit grouping and right group of seeds.
|
Authors: J.H. Kirkbride, Jr., C.R. Gunn, A.L. Weitzman, M.J. Dallwitz, K.R. Thiele
Content last updated April 2003
Released April 2024 on idtools.org



{kind=link}
{kind=link}