Subfamily: Caesalpinioideae.
Phylogenetic Number: 1.4.63.
Tribe: Detarieae.
Group: Berlinia.
Species Studied - Species in Genus: 6 studied; 10 in genus.
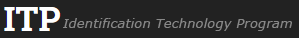
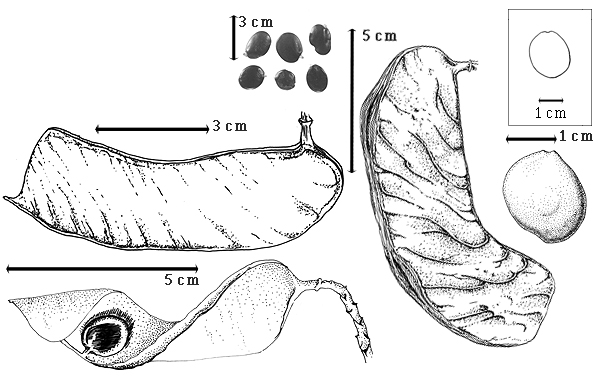
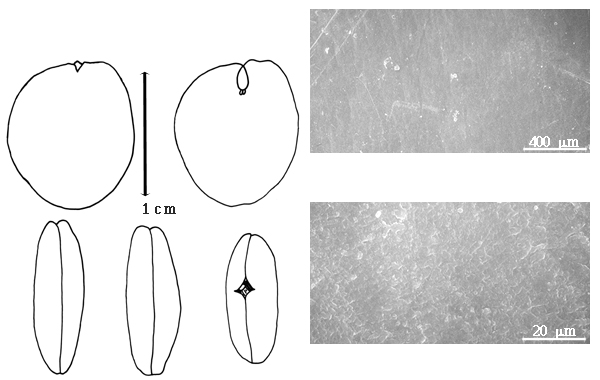
Fruit: A legume; unilocular; 5–26 cm long; 1.8–8 cm wide; 0.5–1 cm thick; 2–9 times longer than wide; with deciduous androecial sheath; with deciduous corolla; with deciduous calyx; without orifice formed by curving of fruit or fruit segments; straight to curved; not plicate; not twisted; symmetrical, or asymmetrical; oblong, or falcate; with both sutures parallelly curved; not inflated; compressed; without beak; tapered at apex; oblique with longitudinal axis of fruit to right-angled with longitudinal axis of fruit; tapered at base; oblique with longitudinal axis of fruit to right angled with longitudinal axis of fruit; with the apex and base uniform in texture; ligneous; seed chambers externally invisible; margin not constricted; margin without sulcus; margin embellished; margin with flange(s), or ridge(s) (up to 3 mm wide); wing(s) absent; stipitate to substipitate; with the stipe 6–7 mm long; with all layers dehiscing; splitting along suture(s). Dehiscence of valves along both sutures; active; with valves elastically and separately enrolling. Replum invisible. Epicarp dull; monochrome; brown; with surface texture uniform; densely velvety brown pubescent and indurate, or glabrous; with hairs erect; eglandular; without spines; not smooth; with elevated features; veined; obliquely veined relative to fruit length and transversely veined relative to fruit length; not tuberculate; not exfoliating; without cracks; without embedded tissue, much thicker than epicarp, running from base to apex. Mesocarp present; surface not veined; 1-layered; without balsamic vesicles; without fibers; without reniform canals; solid; ligneous. Endocarp present; visible; dull to glossy; opaque; monochrome; reddish brown; smooth and spongy (smooth under seeds, spongy around seeds); without adhering pieces of testa; subseptate; not exfoliating; remaining fused to mesocarp and epicarp; entire. Seed(s) 1–5; length oblique to fruit length; neither overlapping nor touching; in 1 series. Funiculus 0.1–1.5 mm long; of 1 length only; flattened; straight. Aril present; dry (?); when dry annular; entire; covering less than 1/2 of seed (?).
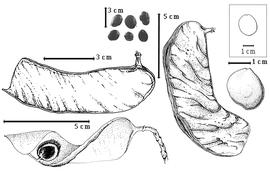
Seed: 12- 13–50 mm long; 0.8–50 mm wide; 3–7 mm thick; not overgrown; angular, or not angular; symmetrical; ovate to oblong, or rectangular; flattened; with surface smooth; without visible radicle and cotyledon lobes; without hilar sinus; without umbo on seed faces; without medial ridge on each face. Cuticle not exfoliating; not inflated; not wrinkled. Testa present; without pieces of adhering epicarp; not adhering to endocarp; free from endocarp; glossy; not modified by a bloom; colored; monochrome; dark brown; glabrous; smooth; osseous. Pleurogram absent. Pseudopleurogram absent. Fracture lines absent. Rim absent. Wing(s) absent. Raphe not visible. Hilum present; fully concealed; concealed by funicular remnant; without faboid split; larger than punctiform; 0.1–2 mm long; with curved outline; elliptic; apical at apex of radicle tip; flush; not within corona, halo, or rim. Lens not discernible. Endosperm absent. Cotyledons smooth; both outer faces convex; both the same thickness; both more or less of equal length; not folded; margin entire 180 degrees from base of radicle; similar at apex; partially concealing radicle; with lobes; with lobes overlapping; with basal groin formed by lobes; with the interface division terminating at base of radicle; without margins recessed; inner face flat; glabrous on inner face. Embryonic axis slightly deflexed (and marginal); perpendicular to length of seed; without a joint evident between the radicle and the cotyledons. Radicle differentiated from cotyledon; centered between cotyledons. Plumule well developed; glabrous.
Julbernardia seeds are unique in the subfamily because of the presence of a rim-aril, slightly deflexed embryonic axis (see also Brenierea, 1.3.05), and groined cotyledon radicle junction. These characters usually are found in seeds from the subfamily Faboideae. Julbernardia seeds do not have a hilar split, a unique character in the legumes for the subfamily Faboideae. At least one species of Anthonotha (1.4.71), A. cladanthea, has seeds with a rim-aril. In 1995, F.J. Breteler informed G.P. Lewis (Corby and Kirkbride, 2002) that Thylacanthus (1.4.49), thought to be a montypic Brazilian genus based on a single collection, is actually J. paniculata (Benth.) Troupin from Mozambique.
Authors: J.H. Kirkbride, Jr., C.R. Gunn, A.L. Weitzman, M.J. Dallwitz, K.R. Thiele
Content last updated April 2003
Released April 2024 on idtools.org

{kind=link}
{kind=link}