Subfamily: Caesalpinioideae.
Phylogenetic Number: 1.1.28.
Tribe: Caesalpinieae.
Group: Caesalpinia.
Species Studied - Species in Genus: 3 studied; 3 in genus.
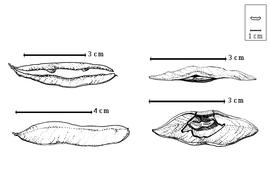
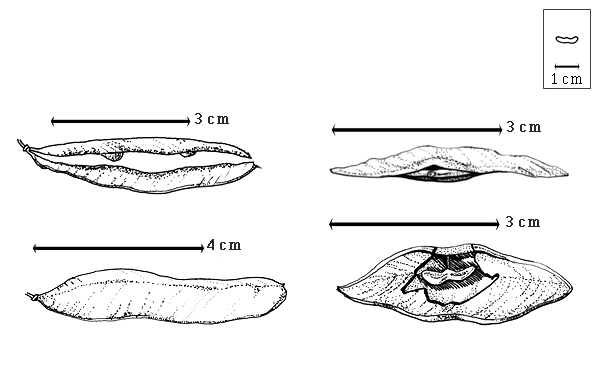
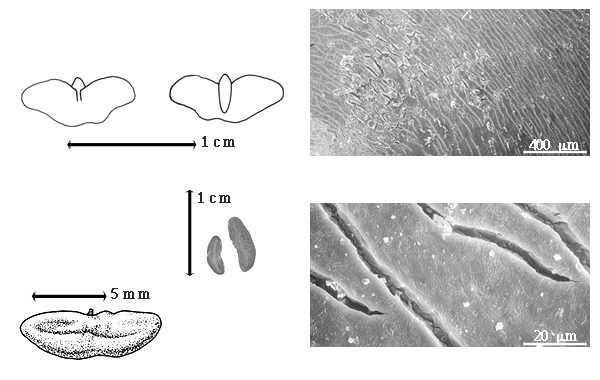
Fruit: A legume; unilocular; 2–4.5 cm long; 0.8–1.2 cm wide; 0.1 cm thick; 2–9 times longer than wide; with deciduous androecial sheath; with deciduous corolla; with deciduous calyx; without orifice formed by curving of fruit or fruit segments; straight to curved; not plicate; not twisted; asymmetrical; lanceolate, or oblong; not inflated; compressed; without beak; long tapered at apex to tapered at apex; aligned with longitudinal axis of fruit; short tapered at base to rounded at base; aligned with longitudinal axis of fruit; with the apex and base uniform in texture; coriaceous; seed chambers externally invisible, or visible; margin constricted, or not constricted (not constricted in 1-seeded fruits); margin slightly constricted along both margins; margin without sulcus; margin plain; wing(s) absent; substipitate; with all layers dehiscing; splitting along suture(s). Dehiscence of valves apical and down to basal and up (along facial line and not the suture). Replum invisible. Epicarp dull; monochrome; reddish brown to purple; with surface texture uniform; glabrous, or glabrate (glabrous in American spp.); with hairs erect; glandular; with glandular hairs (in African spp.); without spines; not smooth; with elevated features; veined; reticulately veined; not tuberculate; not exfoliating; without cracks; without embedded tissue, much thicker than epicarp, running from base to apex. Mesocarp present; surface not veined; 1-layered; without balsamic vesicles; without fibers; without reniform canals; solid; coriaceous. Endocarp present; visible; dull; opaque; monochrome; brown, or tan; without adhering pieces of testa; nonseptate; not exfoliating; remaining fused to mesocarp and epicarp; entire. Seed(s) 1–3; length parallel with fruit length; neither overlapping nor touching; in 1 series. Funiculus 0.1–4 mm long; of 1 length only; thick; straight, or curved (sharply bent). Aril absent.
Britton and Rose (1930) correctly noted that the fruits were "not dehiscent along the sutures, but splitting through the middle of the valves." This character is unique in the subfamily, and perhaps in the family. They as well as Watson and Dallwitz (1983) incorrectly stated that seeds have no endosperm. Polhill and Vidal (1981) noted that there are three species, two in tropical America and the West Indies and one species in Namibia. They did not name which two New World species were recognized and which one was from Africa. Except for data and drawings in Ross (1977b), the African fruits and seeds were not studied.
Authors: J.H. Kirkbride, Jr., C.R. Gunn, A.L. Weitzman, M.J. Dallwitz, K.R. Thiele
Content last updated April 2003
Released April 2024 on idtools.org


{kind=link}
{kind=link}