Subfamily: Faboideae.
Phylogenetic Number: 3.17.05.
Tribe: Carmichaelieae.
Species Studied - Species in Genus: 7 studied; 18 in genus.
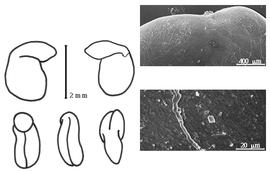
Fruit: A legume; unilocular; 0.5–5 cm long; 0.15–0.8 cm wide; 0.1–0.15 cm thick; 2–9 times longer than wide; with deciduous androecial sheath; with deciduous corolla; with deciduous calyx; without orifice formed by curving of fruit or fruit segments; straight; not plicate; not twisted; asymmetrical, or symmetrical; (sub-) circular, or oblong, or ovate, or elliptic, or linear, or falcate; with both sutures parallelly curved; not inflated; compressed to terete; with beak; straight; with solid beak the same color and texture as fruit; abruptly long acuminate at apex; aligned with longitudinal axis of fruit; rounded at base; aligned with longitudinal axis of fruit; with the apex and base uniform in texture; ligneous; seed chambers externally visible; seed chambers with the raised seed chambers not torulose; margin not constricted; margin without sulcus; margin embellished; margin with thickened sutural areas; wing(s) absent; substipitate; with all layers dehiscing, or indehiscent; splitting along suture(s). Dehiscence of valves along 1 suture; basal and up; active; with valves twisting. Replum visible, or invisible. Epicarp dull; monochrome; black, or brown, or tan; with surface texture uniform; glabrous; without spines; not smooth; with elevated features; veined; transversely veined relative to fruit length; not tuberculate; wrinkled; not exfoliating; without cracks; without embedded tissue, much thicker than epicarp, running from base to apex. Mesocarp present; thin; 1-layered; without balsamic vesicles; without fibers; without reniform canals; solid; ligneous. Endocarp present; visible; dull; opaque; monochrome; brown; fibrous; without adhering pieces of testa; subseptate, or nonseptate; with septa thin (tissue paper-like), flexible; with septa eglandular; chartaceous; not exfoliating; remaining fused to mesocarp and epicarp; without wings. Seed(s) 1–12; length parallel with fruit length; overlapping, or neither overlapping nor touching; in 1 series. Funiculus 0.3–2.5 mm long; of 1 length only; filiform, or thick; straight, or S-curved, or hooked, or plicate. Aril present; dry; when dry rim-aril; entire; brown.
Seed: 2–4.5 mm long; 2–3 mm wide; 1.5–3 mm thick; not overgrown; angular (because of pressure of adjacent seeds in fruit), or not angular; symmetrical, or asymmetrical; irregular, or mitaform, or oblong, or reniform; terete, or compressed; with visible radicle and cotyledon lobes, or without visible radicle and cotyledon lobes; without external groove between radicle and cotyledon lobes; without hilar sinus; without umbo on seed faces; without medial ridge on each face. Cuticle not exfoliating; not inflated; not wrinkled. Testa present, or absent; without pieces of adhering epicarp; not adhering to endocarp; free from endocarp; glossy; not modified by a bloom; colored; monochrome, or mottled and streaked; with frequent mottles, or infrequent mottles; with frequent streaks, or infrequent streaks; black (to bluish-black), or brown, or green (pale), or orange (reddish), or red, or tan, or white (to bluish-white), or yellow, or blue; with black overlay (to bluish-black); glabrous; smooth; coriaceous. Pleurogram absent. Pseudopleurogram absent. Fracture lines absent. Rim absent. Raphe not visible. Hilum present; visible, or fully concealed; concealed by funicular remnant; with faboid split; with the lips of the faboid split the same color as the rest of the hilum; larger than punctiform; 0.3–7 mm long; with straight outline; oblong; marginal according to radicle tip, or between cotyledon and radicle lobe; recessed; not within corona, halo, or rim. Lens discernible, or not discernible; with margins curved; punctiform; not in groove of raphe; adjacent to hilum; 0.5 mm from hilum; mounded, or flush; same color as testa; darker than testa; black, or red (dish); not within corona, halo, or rim. Endosperm present; thick; not pluglike and not resembling tip of radicle; covering entire embryo; adnate to testa, or embryo. Cotyledons smooth; both outer faces convex; both the same thickness; both more or less of equal length; not folded; margin entire 180 degrees from base of radicle; differing at apex (1 concealed by overarching radicle and other auriculate and concealing radicle); not concealing radicle; entire over radicle; without lobes; with the interface division terminating at base of radicle; without margins recessed; white, or red; inner face flat; glabrous on inner face. Embryonic axis deflexed; perpendicular to length of seed; without a joint evident between the radicle and the cotyledons. Radicle differentiated from cotyledon; bulbose; lobe tip curved; with 180 degree turn, or with 90 degree turn; centered between cotyledons; 1/2 to nearly length of cotyledons, or equaling length of cotyledons. Plumule rudimentary; glabrous.>
Simpson (1945) revised the genus recognizing eight subgenera and 41 species based on whether pods dehisce or not and their dehiscence mechanisms, and presented a key to the subgenera essentially using pod characters. Allan (1961) used pod characters as primary divisions in his New Zealand species key. Heenan (1995, 1996, 1998a, 1998c) revised the genus for New Zealand, and carried out cladistic analyses of Carmiachaelia and related genera. He concluded that "Carmichaelia is paraphyletic if Chordospartium (17.03), Corollospartium (17.04), and Notospartium (17.02) are excluded." Therefore he (Heenan, 1998a) merged the four genera, and created new combinations in Carmichaelia for the species of Chordospartium, Corollospartium, and Notospartium. We have chosen to use Polhill's (1981i, 1994b) generic delimitations pending the expression of a general consensus of opinion concerning Heenan's work by taxonomic botanists. Although we studied seeds and fruits of 7 species, the literature cited above provided brief descriptions of seeds and fruits for all species. Heenan's (1995, 1996, 1998a) species count was used.
Tribe Carmichaelieae
Hutchinson (1964) established tribe Carmichaelieae, and Polhill (1981i, 1994a, 1994b) accepted it. Heenan (1995, 1998c), utilizing unpublished nuclear ribosomal DNA ITS data, concluded that "Carmichaelia (17.05) is nested within [the] 'Astragalean clade' of Galegeae" and is the sister group of Clianthus (16.01). He therefore supported the proposal of Sanderson and Wojciehowski (1996) that Carmichaelieae should not be recognized at tribal level, but rather included in Galegeae (16). He (Heenan, 1998c) also carried out cladistic analyses of Carmichaelia (17.05), Chordospartium (17.03), Corollospartium (17.04), and Notospartium (17.02) using morphological and anatomical characters. He (Heenan, 1998a, 1998c) concluded that "Carmichaelia is paraphyletic with Chordospartium, Corollospartium, and Notospartium excluded," and reunited them with Carmichaelia.
Authors: J.H. Kirkbride, Jr., C.R. Gunn, A.L. Weitzman, M.J. Dallwitz, K.R. Thiele
Content last updated April 2003
Released April 2024 on idtools.org


{kind=link}
{kind=link}