Subfamily: Faboideae.
Phylogenetic Number: 3.1.01A.
Tribe: Swartzieae.
Group: Swartzia.
Species Studied - Species in Genus: 2 studied; 2 in genus.
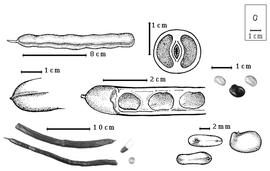
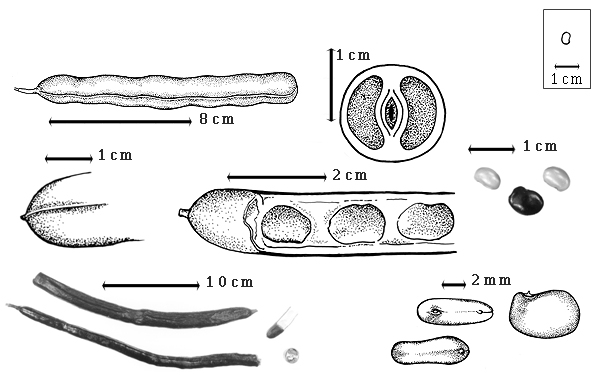
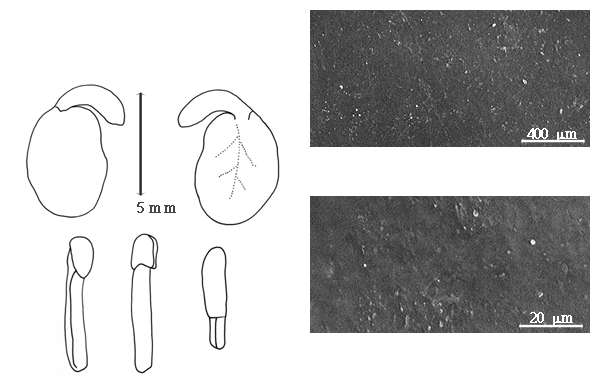
Fruit: A legume; unilocular; 6–30 cm long; 1.3–1.8 cm wide; 1.2–1.6 cm thick; more than 9 times longer than wide; with deciduous androecial sheath; with deciduous corolla; with deciduous calyx; without orifice formed by curving of fruit or fruit segments; straight, or curved (to slightly so), or S-curved; not plicate; not twisted; symmetrical; linear; not inflated; terete; without beak, or with beak (up to 2 mm long); straight; with solid beak the same color and texture as fruit; rounded at apex; aligned with longitudinal axis of fruit; rounded at base, or short tapered at base; aligned with longitudinal axis of fruit; with the apex and base uniform in texture; ligneous; seed chambers externally invisible; margin not constricted; margin with sulcus, or without sulcus; margin plain; wing(s) absent; stipitate; with the stipe 5–10 mm long; indehiscent. Replum invisible. Epicarp dull; monochrome; dark reddish brown; with surface texture uniform; glabrous; eglandular; without spines; not smooth; with elevated features; not veined; not tuberculate; faintly warty; not exfoliating; without cracks; without embedded tissue, much thicker than epicarp, running from base to apex. Mesocarp present; thick; surface not veined; 1-layered (B. fistuloides (H.A.T. Harms) J.H. Kirkbride & J.H. Wiersema), or 2-layered (B. madagascariensis); without balsamic vesicles; without fibers; with reniform canals; with empty space (with or without spongy tissue) within solid layer; ligneous. Endocarp present; visible; glossy; opaque; monochrome; tan; nearly smooth; without adhering pieces of testa; septate; with septa thicker than paper, firm (and mesocarp tissue); with septa eglandular; chartaceous; not exfoliating; remaining fused to mesocarp and epicarp; without wings; entire. Seed(s) 4–20 (determined from fruit length); length parallel with fruit length; neither overlapping nor touching; in 1 series. Funiculus of 1 length only; assumed triangular; straight. Aril absent, or present; dry; when dry tiny rim-aril; entire; tan.
Seed: 6–8 mm long; 4–6 mm wide; 3–3.5 mm thick; not overgrown; not angular; asymmetrical; reniform, or oblong; compressed; with surface smooth; without visible radicle and cotyledon lobes; without hilar sinus; without umbo on seed faces; without medial ridge on each face. Cuticle not exfoliating; not inflated; not wrinkled. Testa present; without pieces of adhering epicarp; not adhering to endocarp; free from endocarp; glossy; not modified by a bloom; colored; monochrome; tan, or brown (dark reddish); glabrous; smooth; coriaceous. Pleurogram absent. Pseudopleurogram absent. Fracture lines absent. Rim absent. Wing(s) absent. Raphe visible; from hilum through lens to base of seed and terminating; not bifurcating; darker than testa; reddish brown; flush. Hilum present; partially concealed, or fully concealed; concealed by aril; with faboid split; with the lips of the faboid split the same color as the rest of the hilum; larger than punctiform; 0.5 mm long; with curved outline; circular; between cotyledon and radicle lobe; recessed; within rim; rim color of testa, or darker than testa. Lens discernible; 1 mm long; with margins curved; elliptic; not in groove of raphe; adjacent to hilum; 0.1–1.7 mm from hilum; mounded; dissimilar color from testa; darker than testa; reddish brown; not within corona, halo, or rim. Endosperm present; thick; not pluglike and not resembling tip of radicle; covering entire embryo; adnate to embryo. Cotyledons smooth; both outer faces convex; both the same thickness; both more or less of equal length; not folded; margin entire 180 degrees from base of radicle; similar at apex; not concealing radicle; entire over radicle; without lobes; with the interface division terminating at base of radicle; without margins recessed; tan; inner face flat; glabrous on inner face. Embryonic axis deflexed; oblique to length of seed; without a joint evident between the radicle and the cotyledons. Radicle differentiated from cotyledon; bulbose; lobe tip curved; deflexed and parallel to cotyledon width; centered between cotyledons; 1/2 to nearly length of cotyledons. Plumule rudimentary; glabrous.
Bobgunnia fistuloides (H.A.T. Harms) J.H. Kirkbride & J.H. Wiersema and B. madagascariensis have been placed in Swartzia, but their seeds and fruits are so incompatible with Swartzia that they were moved to a new genus as a result of these studies (Kirkbride and Wiersema 1997). These species are clearly members of subfamily Faboideae.
Tribe Swartzieae
Swartzieae has been assigned to Caesalpinioideae (Cowan, 1968), Swartzioideae (de Candolle, 1825a, 1825b; Corner, 1976), and Faboideae (Bentham, 1865, Hutchinson, 1964, Cowan, 1981). In 1968, Cowan (1968) was unable "finally to resolve the sub-familial relationship of Swartzia," but in 1981, he (Cowan 1981) placed it in the Faboideae and stated, "... features appear to support the arrangement adopted here (Cowan 1981) with the Swartzieae representing a relatively less-advanced position in the Papilionoideae (Faboideae). This conclusion is now supported by wood anatomy ..., by nodulation proclivity ..., and by chemistry ...; chromosome numbers of n=8, 10 or 14 ..., as well as pollen morphology ..., do not negate this conclusion." In the most recent assessment of the Fabaceae, Polhill (1994a, 1994b) maintained Swartzieae as a basal tribe of Faboideae, "transitional to the Caesalpinioideae." He transferred four genera from Sophoreae (2) into the Swartzieae, Amburana (3.1.15), Ateleia (3.1.13), Cyathostegia (3.1.14), and Holocalyx (3.1.12), and arranged the genera in four groups corresponding to clades in Herendeen's (1994) cladistic analysis. Herendeen carried out cladistic analyses using morphological characters of all Swartzieae genera, 19 genera of Sophoreae, and three Caesalpinioideae genera. He concluded that Swartzieae is polyphyletic and that it should be disbanded and its genera transferred to Sophoreae. Preliminary rbcL data (Doyle et al. 1997) supported his conclusions. Ireland et al. (2000) also carried out molecular phylogenetic studies. They also concluded that Swartizeae is polyphyletic, and suggested that possibly tribe Swartzieae could be maintained with Swartzia (3.1.01), Bobgunnia (3.1.01A), Bocoa (3.1.02), Ateleia (3.1.13), Cyathostegia (3.1.14) and the current remaining Swartzieae genera transferred to other tribes. Our seed data neither support nor refute the overall outlines of Herendeen's cladograms; they are discussed below for a few genera. Ferguson and Skvarla (1991) reported on the pollen morphology of Aldina and Swartzia (1.01), and the nine other genera of Swartzieae are covered in Ferguson and Skvarla (1988). Their data are summarized in a computer-generated key in Vezey et al. (1991). The pollen data for the tribe should be compared with our seed-fruit morphological data.
Authors: J.H. Kirkbride, Jr., C.R. Gunn, A.L. Weitzman, M.J. Dallwitz, K.R. Thiele
Content last updated April 2003
Released April 2024 on idtools.org

{kind=link}
{kind=link}