Subfamily: Faboideae.
Phylogenetic Number: 3.14.26.
Tribe: Aeschynomeneae.
Subtribe: Stylosanthinae.
Species Studied - Species in Genus: 18 studied; 71 in genus.
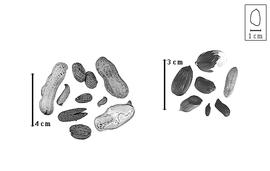
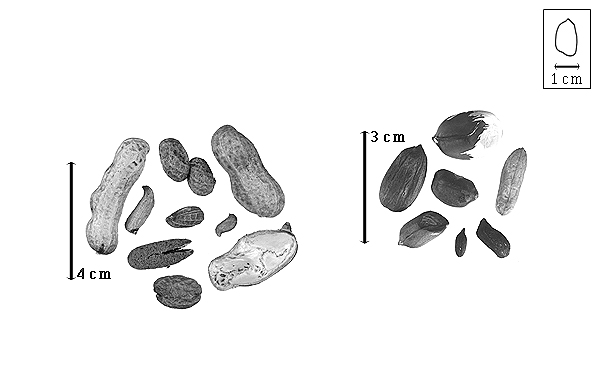
Fruit: A legume; unilocular; 0.8–6 cm long; 0.5–2.1 cm wide; 0.5–2.1 cm thick; 2–9 times longer than wide; with deciduous androecial sheath; with deciduous corolla; with deciduous calyx; without orifice formed by curving of fruit or fruit segments; straight; not plicate; not twisted; asymmetrical, or symmetrical; oblong, or ovate, or irregular; with both sutures unequally curved; not inflated; terete; without beak; rounded at apex; oblique with longitudinal axis of fruit, or right-angled with longitudinal axis of fruit; rounded at base; oblique with longitudinal axis of fruit; with the apex and base uniform in texture; coriaceous; seed chambers externally visible, or invisible; seed chambers with the raised seed chambers not torulose; margin constricted, or not constricted; margin constricted along both margins; margin without sulcus; margin plain; nonstipitate; indehiscent. Replum invisible. Epicarp dull; monochrome; tan; with surface texture uniform; glabrous; eglandular; without spines; not smooth; with elevated features; veined; reticulately veined; not tuberculate; not exfoliating; without cracks; without embedded tissue, much thicker than epicarp, running from base to apex. Mesocarp present; thick; surface uniformly veined; 1-layered; without balsamic vesicles; without fibers; without reniform canals; firm-walled open empty cells; coriaceous. Endocarp present; visible; dull; opaque; monochrome; white; cobwebby, or scurfy; without adhering pieces of testa; nonseptate; chartaceous; not exfoliating; remaining fused to mesocarp and epicarp; entire. Seed(s) 1–3; length parallel with fruit length; touching; in 1 series. Funiculus of 1 length only; thick; straight. Aril absent.
Seed: 7–25 mm long; 4–15 mm wide; 4–15 mm thick; overgrown, 1 seed filling entire fruit cavity, or not overgrown; angular, or not angular; asymmetrical; elliptic, or oblong, or ovate; terete; with visible radicle and cotyledon lobes; without external groove between radicle and cotyledon lobes; without hilar sinus; without umbo on seed faces; without medial ridge on each face. Cuticle not exfoliating; not inflated; not wrinkled. Testa present; without pieces of adhering epicarp; not adhering to endocarp; free from endocarp; dull; not modified by a bloom; colored; monochrome, or bichrome (reddish-brown and white); reddish brown, or white; glabrous; smooth, or not smooth; with elevated features; veined; chartaceous. Pleurogram absent. Pseudopleurogram absent. Fracture lines absent. Rim absent. Raphe not visible. Hilum present; fully concealed; concealed by funicular remnant; without faboid split; larger than punctiform; 2 mm long; with curved outline; elliptic; subapical to radicle tip; flush; not within corona, halo, or rim. Lens not discernible. Endosperm absent, or present (according to Kubicek (1970) and see Notes); trace; not pluglike and not resembling tip of radicle; covering entire embryo; adnate to testa. Cotyledons smooth; both outer faces convex; both the same thickness; both more or less of equal length; not folded; margin entire 180 degrees from base of radicle; similar at apex; partially concealing radicle; split over radicle; with lobes; with lobes not touching; with the interface division terminating at base of radicle; without margins recessed; tan; inner face flat; glabrous on inner face. Embryonic axis straight; parallel to length of seed; without a joint evident between the radicle and the cotyledons. Radicle differentiated from cotyledon; bulbose; lobe tip straight, or curved; straight with embryonic axis; centered between cotyledons; less than 1/2 length of cotyledons. Plumule well developed; glabrous.
Hoehne (1940) and Hermann (1954) monographed the genus, and Resslar (1980) reviewed the nomenclature. Rudd (1981a) used the same number of recognized species as Resslar, and both noted that possibly 40 (Rudd) to 70 (Resslar) species were undescribed. Wynne and Halward (1989) recognized 33 species including several nomum nuda species names. Krapovickas and Gregory (1994) monographed Arachis recognizing 69 species in the genus. José F.M. Valls (pers. commun., 1994) has informed us that several additional new species of Arachis have been discovered. Kubicek (1970) stated that "contrary to reports by other investigators the endosperm appeared to be present as a single layer which covers the cotyledons in mature seeds." Gregory and Gregory (1979) studied interspecific hybrids. Bentham (1841) compared the structures and affinities of Arachis and Voandzeia (now Vigna 10.65). Charles F. Simpson, USDA/ARS, made a major contribution of seeds and fruits to BARC.
Tribe Aeschynomeneae
Rudd (1981a) recognized four subtribes of Aeschynomeneae: Ormocarpinae V.E. Rudd (genera 3.14.01–3.14.08), Aeschynomeninae (genera 3.14.09–3.14.16), Discolobinae (A.E. Burkart) V.E. Rudd (genus 3.14.17: Discolobium), Poiretiinae (A.E. Burkart) V.E. Rudd (genera 3.14.18–3.14.21), and Stylosanthinae (G. Bentham) V.E. Rudd (genera 3.14.22–13.4.26). Tribal and subtribal placement of Diphysa is based on Lavin (1987; Polhill, 1994a, 1994b), and not on Polhill and Sousa (1981), who placed Diphysa in Robinieae. Bailey et al. (1997), using the chloroplast rpl2 intron and ORF184, suggested that Brya (11.01), Cranocarpus (11.02), Phylacium (11.22), and Neocollettia (11.26) are not members of Desmodieae (11) and that they probably belong in Aeschynomeneae.
Authors: J.H. Kirkbride, Jr., C.R. Gunn, A.L. Weitzman, M.J. Dallwitz, K.R. Thiele
Content last updated April 2003
Released April 2024 on idtools.org


{kind=link}
{kind=link}