Subfamily: Faboideae.
Phylogenetic Number: 3.2.01.
Tribe: Sophoreae.
Group: Myroxylon.
Species Studied - Species in Genus: 6 studied; 16 in genus.
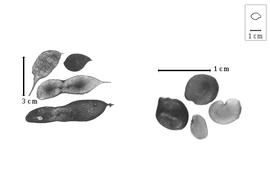
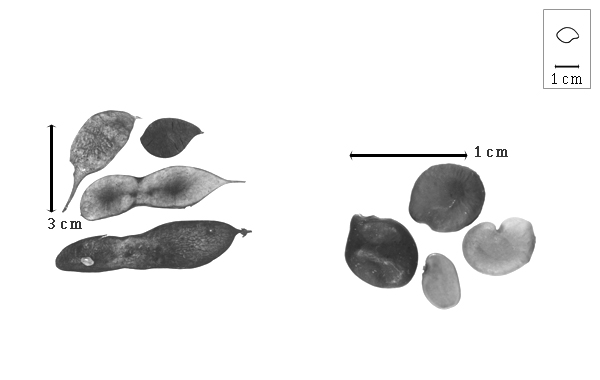
Fruit: A legume, or a nutlet; unilocular; 1.7–9.2 cm long; 1.4–2.4 cm wide; 0.2–0.7 cm thick; length less than twice as long as width, or 2–9 times longer than wide; with deciduous androecial sheath; with persistent calyx, or deciduous calyx; with calyx shorter than fruit; without orifice formed by curving of fruit or fruit segments; straight; not plicate; not twisted; asymmetrical, or symmetrical; elliptic, or moniliform, or irregular; with both sutures unequally curved; not inflated; flattened, or compressed; without beak; short tapered at apex, or rounded at apex; aligned with longitudinal axis of fruit; tapered at base to rounded at base; aligned with longitudinal axis of fruit, or oblique with longitudinal axis of fruit; with the apex and base uniform in texture; coriaceous to ligneous; seed chambers externally visible, or invisible; seed chambers with the raised seed chambers not torulose; margin not constricted, or constricted; margin slightly constricted along both margins; margin without sulcus; margin plain, or embellished; margin with ridge(s) (slight); wing(s) absent; nonstipitate, or substipitate, or stipitate; with the stipe 2–7 mm long; indehiscent. Replum invisible. Epicarp dull to semiglossy; monochrome, or multicolored; bichrome; brown; with brown overlay; with surface texture uniform; glabrous, or glabrate, or pubescent and indurate; with hairs erect; with 1 type of pubescence; puberulent; with pubescence golden; with pubescence uniformly distributed; with simple hairs; pliable; with hair bases plain, or swollen; eglandular, or glandular; with glandular hairs; without spines; not smooth; with elevated features; veined; reticulately veined; not tuberculate; papillose; not exfoliating; with cracks, or without cracks; cracking irregular; without embedded tissue, much thicker than epicarp, running from base to apex. Mesocarp present; thick; surface not veined; 1-layered, or 2-layered; without balsamic vesicles; without fibers; without reniform canals; solid; with spongy layer over solid layer; (sub-) ligneous, or coriaceous, or chartaceous. Endocarp present; visible; dull; opaque; monochrome; brown to yellow; scurfy to smooth, or fibrous; without adhering pieces of testa; nonseptate; chartaceous; not exfoliating; remaining fused to mesocarp and epicarp; without wings; entire. Seed(s) 1–3; length parallel with fruit length, or oblique to fruit length; neither overlapping nor touching; in 1 series. Aril present; dry; when dry rim-aril; fimbriate; ivory.
Seed: 7–11.3 mm long; 5–8 mm wide; 2.5–5.2 mm thick; not overgrown; not angular; symmetrical or nearly symmetrical; ovate, or circular, or oblong, or reniform; compressed to terete; with surface smooth; without visible radicle and cotyledon lobes; without hilar sinus; without umbo on seed faces; without medial ridge on each face. Cuticle not exfoliating; not inflated; not wrinkled. Testa present; without pieces of adhering epicarp; not adhering to endocarp; free from endocarp; dull; not modified by a bloom; colored; monochrome, or mottled; with infrequent mottles; reddish brown to brown; with brown overlay; glabrous; smooth; chartaceous. Pleurogram absent. Pseudopleurogram absent. Fracture lines absent. Rim absent. Wing(s) absent. Raphe visible, or not visible; from hilum through lens and base of seed to point opposite hilum; not bifurcating; darker than testa; brown; flush. Hilum present; partially concealed; concealed by aril; with faboid split; with the lips of the faboid split the same color as the rest of the hilum; larger than punctiform; 0.8–2 mm long; with curved outline; elliptic; subapical to radicle tip; flush, or recessed; within rim, or not within corona, halo, or rim; rim color darker than testa. Lens discernible, or not discernible; 2–3 mm long; with margins straight; linear, or wedge-shaped; not in groove of raphe; confluent with hilum; flush; similar color as testa; darker than testa; brown; not within corona, halo, or rim. Endosperm present; thick, or trace; not pluglike and not resembling tip of radicle; covering entire embryo, or restricted to region of embryo; adnate to testa. Cotyledons smooth; both outer faces convex; both the same thickness; both more or less of equal length; not folded; margin entire 180 degrees from base of radicle; similar at apex; partially concealing radicle; notched at radicle; with lobes; with lobes not touching; without basal groin formed by lobes; with the interface division terminating at base of radicle; without margins recessed; tan; inner face with central ridge on 1 and central groove on other; glabrous on inner face. Embryonic axis straight, or deflexed (slightly); parallel to length of seed; without a joint evident between the radicle and the cotyledons. Radicle differentiated from cotyledon; bulbose; lobe tip straight; straight with embryonic axis, or deflexed and parallel to cotyledon length (slightly); centered between cotyledons; less than 1/2 length of cotyledons. Plumule rudimentary; glabrous.
Southern Mexico to northern Argentina.
New World; Mexico, Central America, and South America; Argentina, Brazil, and Ecuador.
Polhill (1981b) noted that Lovanafia M. Peltier (2.33) is closely related to this genus, but Yakovlev (1977) and Polhill (1994a, 1994b) included Lovanafia in Dicraeopetalum (2.33). Bridgewater and Striton (1997) studied the infraspecific variation of A. dasycarpum (J.R.T. Vogel) G.P. Yakovlev, and concluded that it can not be segrated infraspecifically.
Tribe Sophoreae
Polhill (1981b) stated that the Sophoreae s.l. is a tribe of convenience between the Caesalpinioideae and the bulk of the Papilionoideae, sharply defined from neither. He transferred four genera from Sophoreae into the Swartzieae (1), Amburana (3.1.15), Ateleia (3.1.13), Cyathostegia (3.1.14), and Holocalyx (3.1.12), following Herendeen's (1995) cladistic analyses. Herendeen performed cladistic analyses for all Swartzieae genera, sensu Cowan (1981), 19 Sophoreae genera, and three Caesalpinioideae genera. He concluded that Swartzieae is polyphyletic and that it should be disbanded and its genera transferred to Sophoreae. Preliminary rbcL data (Doyle et al. 1997) supported his conclusions.
Authors: J.H. Kirkbride, Jr., C.R. Gunn, A.L. Weitzman, M.J. Dallwitz, K.R. Thiele
Content last updated April 2003
Released April 2024 on idtools.org


{kind=link}
{kind=link}