Exotic
Thaumatotibia leucotreta (Meyrick) (Tortricidae: Olethreutinae: Grapholitini)
Common names: false codling moth (FCM)
Synonyms: roerigii (Thaumatotibia)
FWL: 7.0-8.0 mm (males); 9.0-10.0 (females)
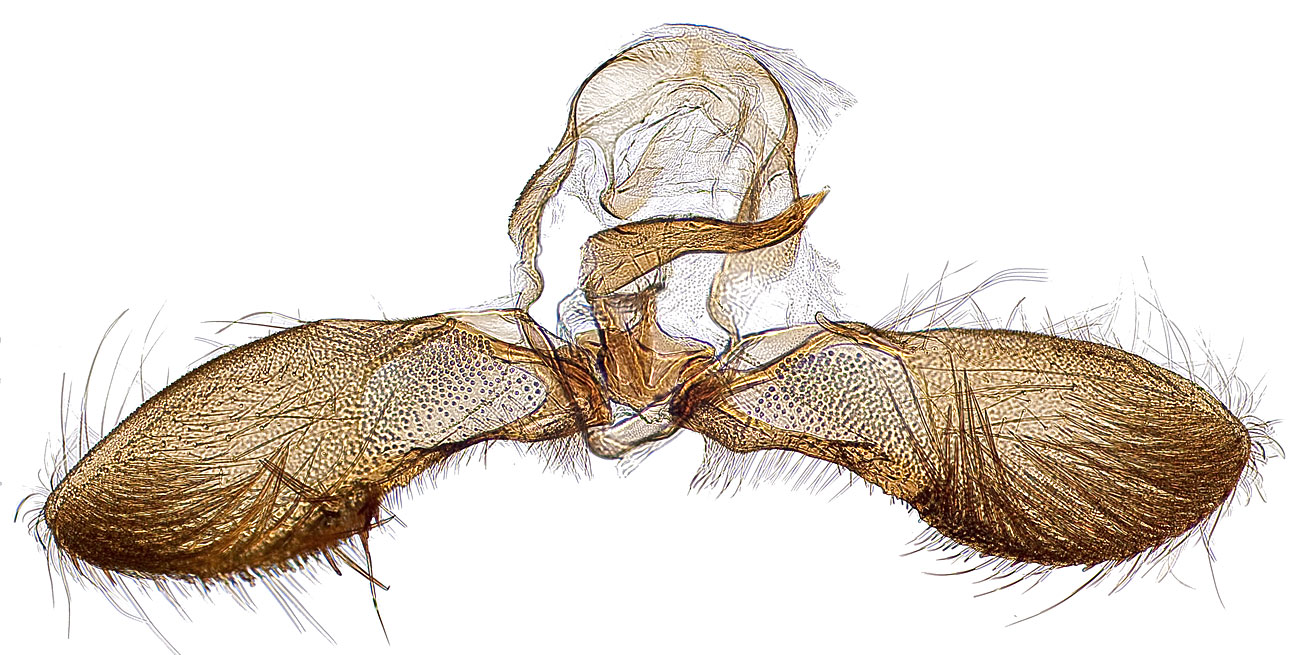
Adults of T. leucotreta are sexually dimorphic, and the two sexes differ in overall size, wing shape, and male secondary sexual characters. Male forewings are triangular with an acute apexapex:
the point furthest removed from the base or at the end of the costal area  , while female forewings are more elongate with a rounded apexapex:
, while female forewings are more elongate with a rounded apexapex:
the point furthest removed from the base or at the end of the costal area . Both sexes exhibit a combination of the same forewing pattern elements: a small white dot near the end of the discal cell; a patch of raised, usually rust or orange colored scales near the middle of the wing; a distinct question mark-shaped band of dark scales along the termentermen:
the outer edge of the forewing  ; and a semicircular band of dark scales in the middle of the costacosta:
; and a semicircular band of dark scales in the middle of the costacosta:
the anterior margin of each wing  . Males are easily distinguished by a semicircular pocket of opalescent scales at the distaldistal:
. Males are easily distinguished by a semicircular pocket of opalescent scales at the distaldistal:
farthest from body, distant from point of attachment end of vein CuA2 on the hindwing, tufts of modified scales on the hind tibia, and an enlargement of the inner apical spur on the hind tibia. Males lack a forewing costal foldforewing costal fold:
end of vein CuA2 on the hindwing, tufts of modified scales on the hind tibia, and an enlargement of the inner apical spur on the hind tibia. Males lack a forewing costal foldforewing costal fold:
a flap or fold at the base of the forewing that contains specialized sex scales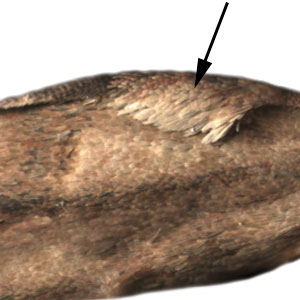 .
.
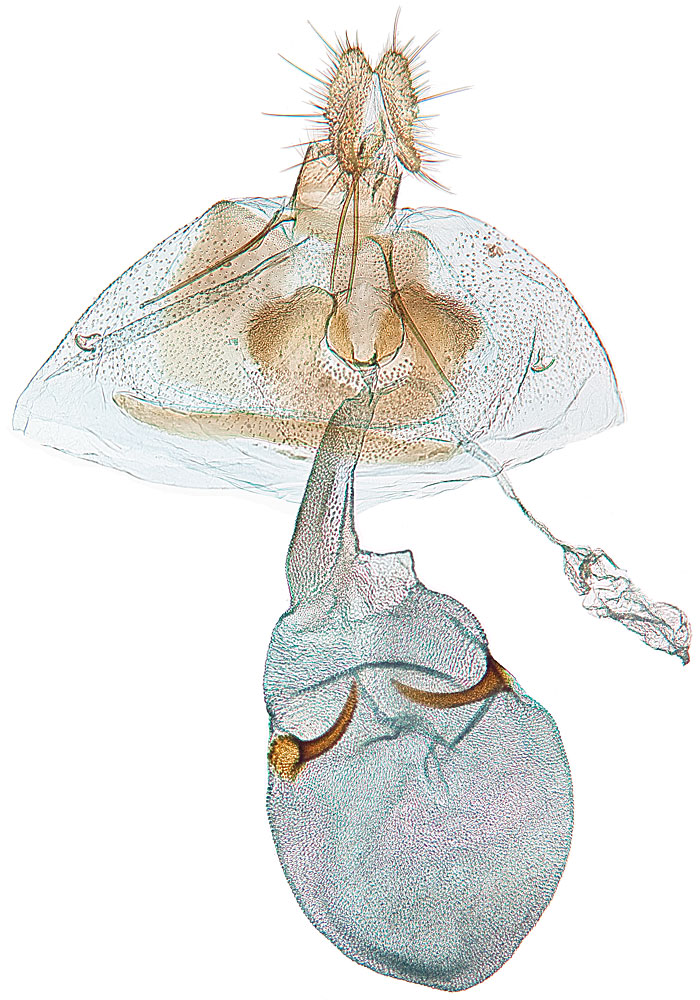
Male genitalia are characterized by a rounded tegumentegumen:
dorsal section of the transverse ring in male genitalia; often heavily sclerotized 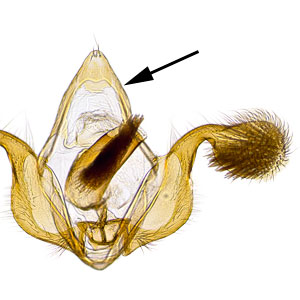 lacking an uncusuncus:
lacking an uncusuncus:
a sclerotized process which is fused to the posterodorsal margin of tergum IX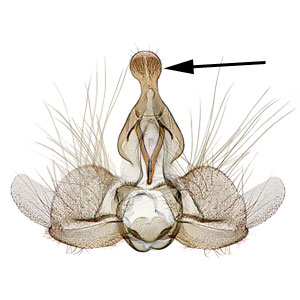 or sociisocii:
or sociisocii:
a pair of lightly sclerotized setose lobes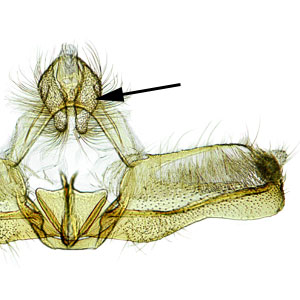 , large rounded valvaevalvae:
, large rounded valvaevalvae:
plural of "valva" , and a tapered aedeagusaedeagus:
, and a tapered aedeagusaedeagus:
the male intromittent organ (penis); see "phallus" 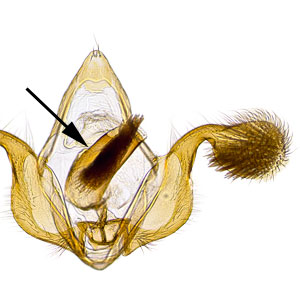 that is upcurved distally. Female genitalia are characterized by a semicircular sterigmasterigma:
that is upcurved distally. Female genitalia are characterized by a semicircular sterigmasterigma:
the sclerotized region surrounding the female ostium bursae 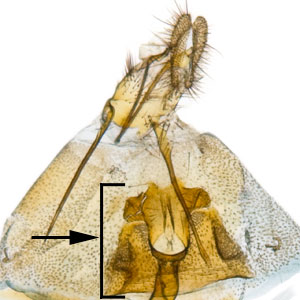 , narrow ductus bursaeductus bursae:
, narrow ductus bursaeductus bursae:
a membranous tube connecting the ostium bursae to the corpus bursae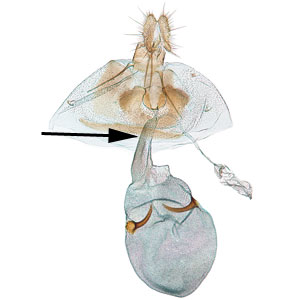 , and large rounded corpus bursaecorpus bursae:
, and large rounded corpus bursaecorpus bursae:
a dilated membranous sac at the anterior end of the bursa copulatrix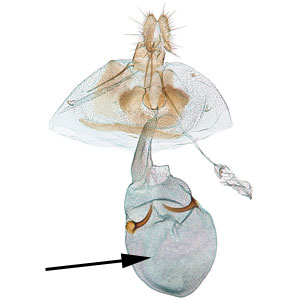 with a pair of thorn-shaped signasigna:
with a pair of thorn-shaped signasigna:
plural of "signum"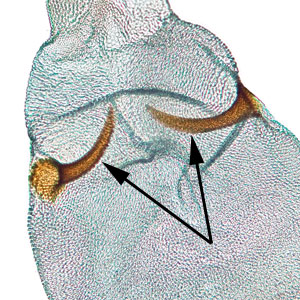 .
.
The following account is summarized from Gilligan et al. (2011c).
Last instar larvae are approximately 12-18 mm long with a yellowish brown to dark brown head and prothoracic shieldprothoracic shield:
a sclerotized plate on the dorsal surface of the prothorax  . The abdomen is orange to pink with large pinaculapinacula:
. The abdomen is orange to pink with large pinaculapinacula:
flattened sclerotized plates on a caterpillar that bear the setae that are darker than body color. See below for a detailed description of larval chaetotaxychaetotaxy:
that are darker than body color. See below for a detailed description of larval chaetotaxychaetotaxy:
the arrangement of setae (in reference to Lepidoptera larvae), often depicted on a "setal map"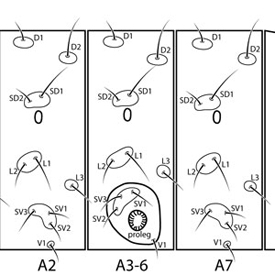 .
.
The semicircular pocket of scales on the hindwing can be used to separate T. leucotreta males from all other North American tortricids. Many species of Cryptophlebia have similar scale pockets, but no species of Cryptophlebia occur in North America. A dissection can be used to confirm female identity.
Thaumatotibia leucotreta larvae can be distinguished from those of many other North American tortricids by the following combination of characters: L pinaculumpinaculum:
singular of "pinacula" on T1 enlarged, extending beneath and beyond (posteradposterad:
on T1 enlarged, extending beneath and beyond (posteradposterad:
towards the posterior  of) spiracle; D1 and SD1 on A9 on same pinaculumpinaculum:
of) spiracle; D1 and SD1 on A9 on same pinaculumpinaculum:
singular of "pinacula", separate from D2; L group on A9 trisetose; anal combanal comb:
a toothed structure on the last abdominal segment used to eject frass away from the feeding larva; also termed "anal fork" 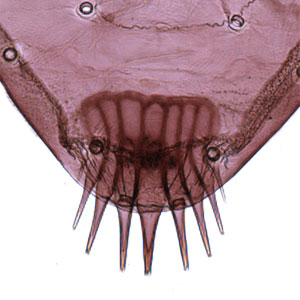 present with 2–10 teeth. Other larval characters include: SD2 on A1–8 highly reduced or appearing absent; SV groups on A1, 2, 7, 8, 9 with 3:3:2:2:1 setae; spiracle on A8 displaced posteradposterad:
present with 2–10 teeth. Other larval characters include: SD2 on A1–8 highly reduced or appearing absent; SV groups on A1, 2, 7, 8, 9 with 3:3:2:2:1 setae; spiracle on A8 displaced posteradposterad:
towards the posterior of SD pinaculumpinaculum:
singular of "pinacula"; D2 setae on A9 on shared mid-dorsal " saddlesaddle:
in reference to the D2 pinacula on larval segment A9; both setae are on a single, fused pinaculum in the mid-dorsal region of the segment  " pinaculumpinaculum:
" pinaculumpinaculum:
singular of "pinacula"; V setae on A9 slightly further apart than those on A8. The anal combanal comb:
a toothed structure on the last abdominal segment used to eject frass away from the feeding larva; also termed "anal fork" may be greatly reduced in some individuals.
Larval characters listed here are not completely diagnostic, and T. leucotreta larvae are difficult to separate from Cochylini larvae, which can share the same set of character states (e.g., enlarged L-pinaculum on A9, spiracle on A8 displaced posteradposterad:
towards the posterior , anal combanal comb:
a toothed structure on the last abdominal segment used to eject frass away from the feeding larva; also termed "anal fork" present). Two possible distinguishing features are SV counts of 3:3:2:2:2 and a bisetose L-group on A9 in many Cochylini.
The following account is summarized from Gilligan et al. (2011c).
Thaumatotibia leucotreta is not known to diapause, and development is continuous with adults present year-round. As many as 10 generations are possible per year in South Africa. The absence of a diapause may lead to host shifts and varied developmental rates in times of drought or when preferred host plants are unavailable.
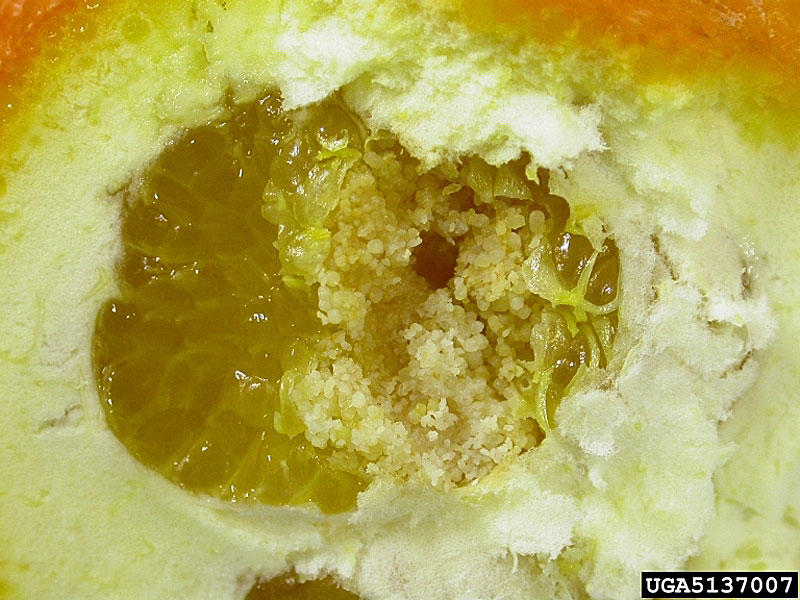
Females deposit eggs singly or in small groups on the surface of smooth fruit. A single female may produce between 87-456 eggs in her lifetime (with a maximum of 799). On fruit, larvae tunnel into the pith or feed beneath the surface. On cotton, larvae mine the wall of the boll and later move into the center of the boll to feed on the seeds. Larvae complete five instars. Last instar larvae exit the fruit or boll, drop from the host plant, and pupate in a silken cocoon in the soil, under leaf litter, or in bark crevices.
Larvae cause significant damage by feeding directly on fruit or bolls. Feeding in citrus fruit can result in premature ripening and fruit drop as well as secondary infection by fungi. Larval feeding in cotton results in secondary infection by fungus and bacteria, causing rotting of the bolls. Feeding in avocado fruit results in lesions on the fruit and secondary infection by bacteria and fungi. Chemical control of this species is difficult due to the highly polyphagous, internal feeding larvae, and crop losses can be as high as 10–20% during serious citrus infestations. Control of false codling moth in South Africa is achieved through a combination of chemical control, mating disruption, attract and kill, natural enemies, and sterile insect technique (SIT).
Larvae of T. leucotreta are highly polyphagous and have been recorded feeding on more than 50 species of plants in over 30 families. In Africa, false codling moth is a serious pest of citrus, cotton, and avocado. It has also been reported causing serious damage to corn, guava, macadamia, mango, peach, and other horticultural crops.
False codling moth is one of the most commonly intercepted tortricids on pepper and eggplant at U.S. ports-of-entry.
| Host plant | Host plant family | Reference(s) |
| Mangifera indica | Anacardiaceae | Grove et al. 2012Grove et al. 2012: Grové, T., De Beer, M.S., Joubert, P.H. 2012 Host status of commercial mango cultivars to Thaumatotibia leucotreta (Lepidoptera: Tortricidae) in South Africa, Journal of Economic Entomology. 105(6): 1954-1962. |
| Sclerocarya birrea | Anacardiaceae | Brain 1929Brain 1929: Brain, C. K. 1929. Insect pests and their control in South Africa. Die Nasionale Pers Beperk, Cape Town. |
| Annona muricata | Annonaceae | Whittle 1984 |
| Annona reticulata | Annonaceae | Whittle 1984 |
| Lettowianthus stellatus | Annonaceae | Brown et al. 2014 |
| Monodora grandidieri | Annonaceae | Brown et al. 2014 |
| Uvaria acuminata | Annonaceae | Brown et al. 2014 |
| Uvaria scheffleri | Annonaceae | Brown et al. 2014 |
| Xylopia parviflora | Annonaceae | Brown et al. 2014 |
| Landolphia sp. | Apocynaceae | Brown et al. 2014 |
| Aristolochia albida | Aristolochiaceae | Brown et al. 2014 |
| Calotropis procera | Asclepiadaceae | Whittle 1984 |
| Ceiba pentandra | Bombacaceae | Stotter 2009Stotter 2009: Stotter, R.L., 2009. Spatial and temporal distribution of false codling moth across landscapes in the Citrusdal area (Western Cape Province, South Africa). MSc Thesis, University of Stellenbosch, 101 pp. Available online: https://scholar.sun.ac.za/items/b2a733dd-5247-4eb0-b40f-32b732064d1a [Accessed: 17 April 2024] |
| Ananas comosus | Bromeliaceae | Whittle 1984 |
| Capparis sp. | Capparaceae | Whittle 1984 |
| Catha edulis | Celastraceae | Whittle 1984 |
| Salacia elegans | Celastraceae | Brown et al. 2014 |
| Salacia leptoclada | Celastraceae | Brown et al. 2014 |
| Hirtella zanzibarica | Chrysobalanaceae | Brown et al. 2014 |
| Garcinia mangostana | Clusiaceae | Stotter 2009Stotter 2009: Stotter, R.L., 2009. Spatial and temporal distribution of false codling moth across landscapes in the Citrusdal area (Western Cape Province, South Africa). MSc Thesis, University of Stellenbosch, 101 pp. Available online: https://scholar.sun.ac.za/items/b2a733dd-5247-4eb0-b40f-32b732064d1a [Accessed: 17 April 2024] |
| Combretum apiculatum | Combretaceae | Whittle 1984 |
| Combretum zeyheri | Combretaceae | Whittle 1984 |
| Agelaea pentagyna | Connaraceae |
View full screen host table here
Thaumatotibia leucotreta is widely distributed across Africa and has been reported from approximately 40 countries on the African continent. It is not considered established outside of Africa although it is commonly intercepted during quarantine inspections in the U.S. and Europe. It was detected in a pheromone trap in Ventura County, California in 2008, although it does not appear to have become established (Gilligan et al. 2011cGilligan et al. 2011c:
Gilligan, T. M., Epstein, M. E., Hoffman, K. M. 2011. Discovery of false codling moth, Thaumatotibia leucotreta (Meyrick), in California (Lepidoptera: Tortricidae). Proceedings of the Entomological Society of Washington. 113: 426-435.).