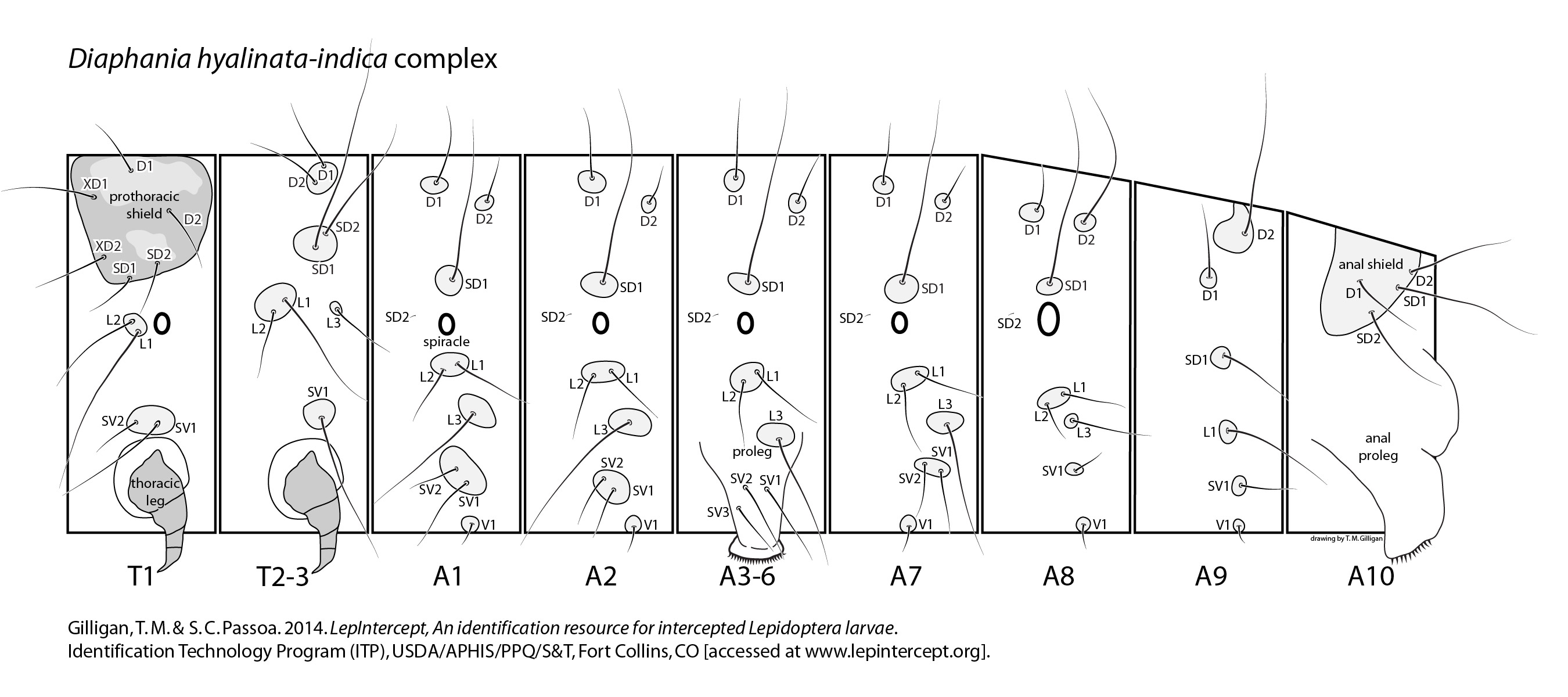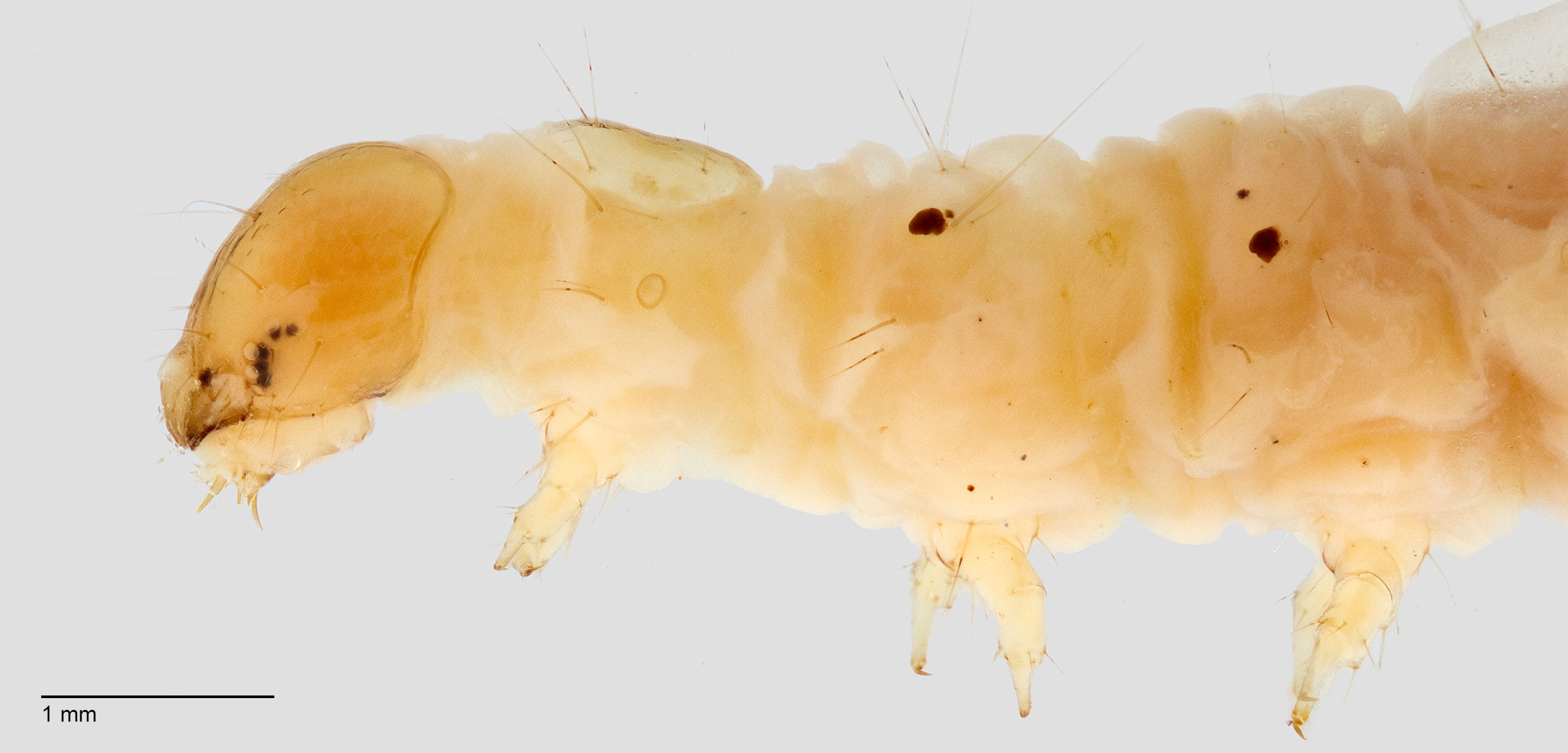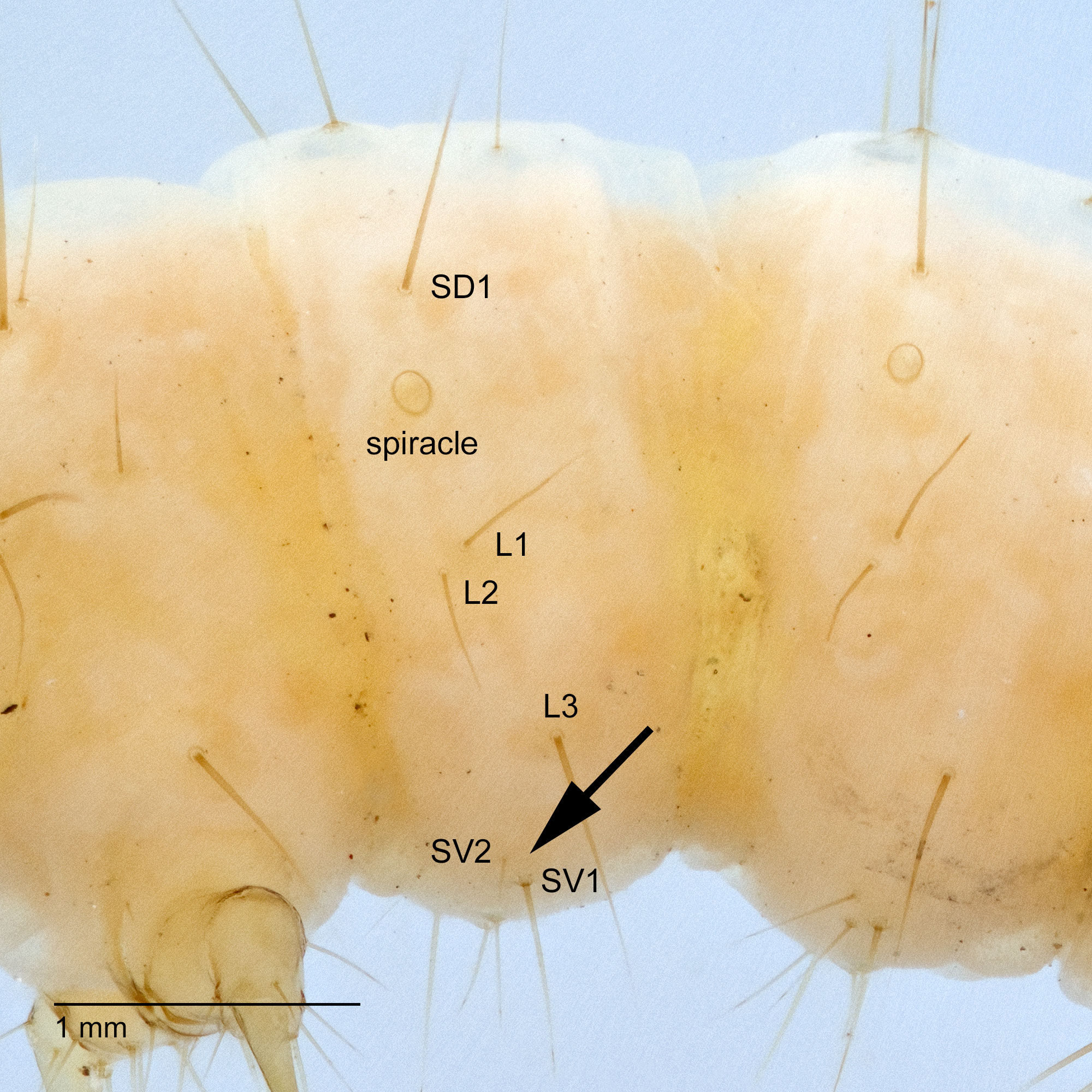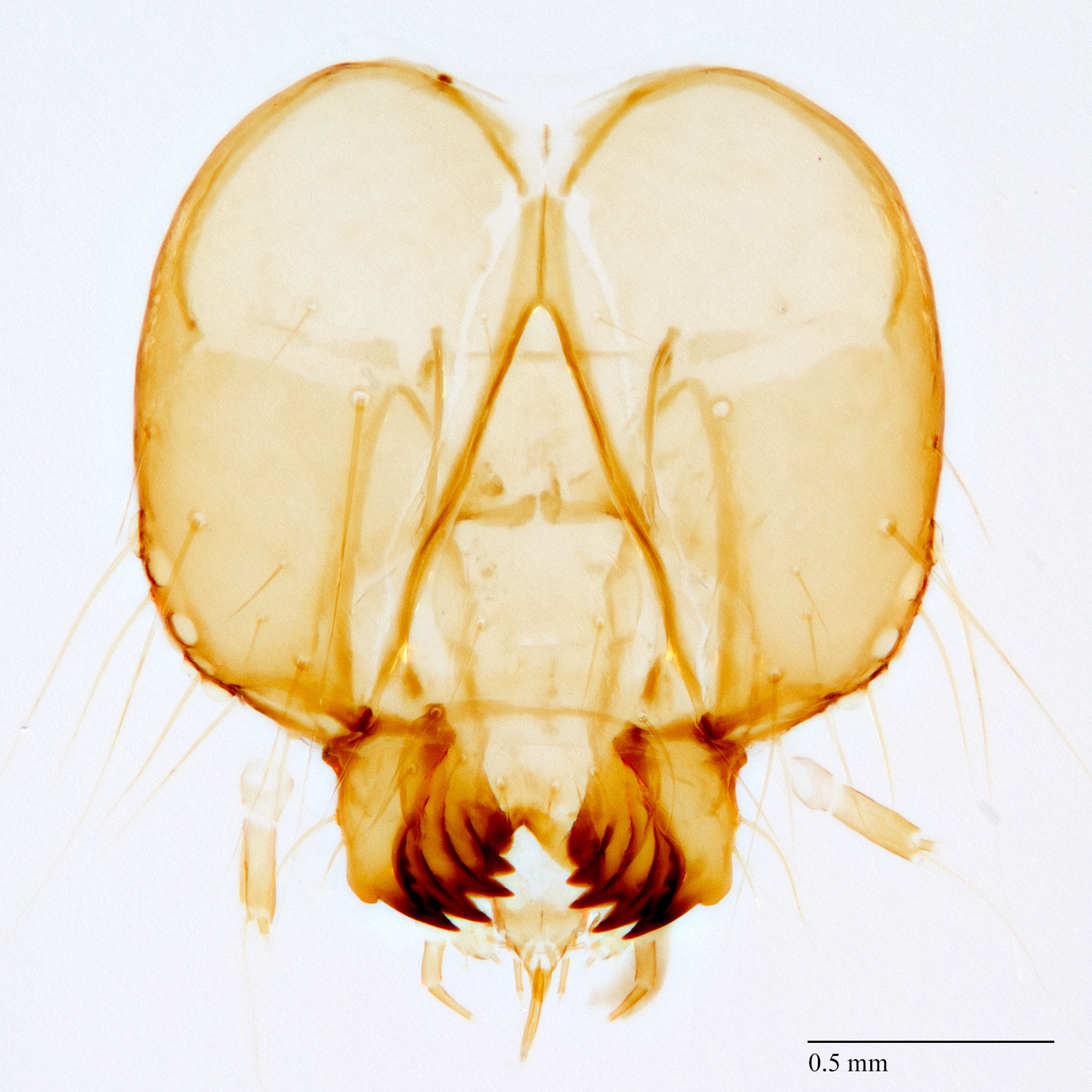
Pyraloidea: Crambidae: Spilomelinae: Diaphania hyalinata-indica complex
[too many to list - see http://www.pyraloidea.org for complete taxonomy]
Larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
in the D. hyalinata-indica complex are most frequently intercepted from Mexico (33%) and the Caribbean (>60%). Common host/origin combinations are listed below:
| Origin | Host(s) |
|---|---|
| Dominican Republic | Luffa, Momordica |
| Haiti | Momordica |
| Jamaica | Momordica |
| Mexico | Cucurbita, Sechium |
Diaphania hylinata occurs from Canada to Argentina. Diaphania indica occurs in Florida, tropical America, and the Old World tropics.
Identification of larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
in the D. hyalinata-indica complex is based on the absence of a genal spot, presence of an outer tooth on the mandible, and the bisetosebisetose:
two setae
SV group on A1. None of these characters are very distinctive and numerous sibling species of Diaphania exist with unknown larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
. These uncertainties all suggest a need for caution; see the Detailed Information tab.
Diaphania indica and D. hyalinata were placed in the D. hyalinata species group by Clavijo (1990)Clavijo (1990):
Clavijo, J. A. 1990. Systematics of black and white species of the genus Diaphania Hubner (1818) (Lepidoptera: Pyralidae: Pyraustinae). Ph.D. Dissertation. McGill University. Montreal, Canada. 215 pp.; later called the D. indica complex by Solis (2011)Solis (2011):
Solis, M. A. 2011. Key to selected Pyraloidea (Lepidoptera) larvae intercepted at U. S. ports of entry: revision of Pyraloidea in "Keys to some frequently intercepted lepidopterous larvae" by Weisman 1986 (updated 2006).. These two species are treated together because the larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
are difficult to separate and there is evidence that both are intercepted at United States ports. There are other species of Diaphania in Latin America that are similar to both D. indica and D. hyalinata in adult coloration (Arias and Clavijo 2001Arias and Clavijo 2001:
Arias, Q. and J. A. Clavijo. 2001. Clave pictorica de las especies de Diaphania Hubner, 1818 (Lepidoptera: Crambidae) de Venezuela. Entomotropica 16: 1-13.) and potentially part of this complex, but at present these other species do not appear to be pest species of concern to APHIS.
The larvalarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of D. hyalinata, called the melonworm, was partially described by Smith (1911)Smith (1911):
Smith, R. I. 1911. Two important cantaloupe pests. North Carolina Agricultural Experimental Station Bulletin 214. 45 pp., Peterson (1962)Peterson (1962):
Peterson, A. 1962. Larvae of insects: an introduction to Nearctic species. Part I: Lepidoptera and plant infesting Hymenoptera. Columbus, Ohio. 315 pp., Weisman (1986)Weisman (1986):
Weisman, D. M. 1986. Keys for the identification of some frequently intercepted lepidopterous larvae. U.S. Department of Agriculture, Animal and Plant Health Inspection Service, Plant Protection and Quarantine series 81-47. 64 pp., Neunzig (1987)Neunzig (1987):
Neunzig, H. H. 1987. Pyralidae (Pyraloidea), pp. 464-494. In F.W. Stehr (ed.). Immature Insects. Kendall Hunt Publishing Company. Dubuque, Iowa., Solis (2011)Solis (2011):
Solis, M. A. 2011. Key to selected Pyraloidea (Lepidoptera) larvae intercepted at U. S. ports of entry: revision of Pyraloidea in "Keys to some frequently intercepted lepidopterous larvae" by Weisman 1986 (updated 2006)., and Schnitzler et al. (2012)Schnitzler et al. (2012):
Schnitzler, F. R., J. W. Haw, L. Kumarasinghe and S. George. 2012. Identification Guide to Lepidoptera Larvae Intercepted on Trade Pathways. Bulletin of the Entomological Society of New Zealand 15. 105 pp.. Color photographs were published by King and Saunders (1984)King and Saunders (1984):
King, A. B. S. and J. L. Saunders. 1984. The invertebrate pests of annual food crops in Central America. Overseas Development Administration. 166 pp., Passoa (1985)Passoa (1985):
Passoa, S. 1985. Taxonomy of the larvae and pupae of economically important Pyralidae in Honduras. Master's Thesis. University of Florida. Gainesville. 486 pp., Schmutterer et al. (1990), Sparks and Liu (2001)Sparks and Liu (2001):
Sparks, A. N. and T.-X. Liu. 2001. A key to common caterpillar pests of vegetables. Texas A&M Agricultural Extension Service. AgriLife Communications and Marketing E-532. 8 pp., and Cranshaw (2004)Cranshaw (2004):
Cranshaw, W. 2004. Garden Insects of North America. Princeton University Press. Princeton, New Jersey. 678 pp..
The larvalarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of D. indica, called the pumpkin caterpillarcaterpillar:
a larva with a conspicuous head, three pairs of thoracic legs, and prolegs; the larva of a butterfly, moth, sawfly, or scorpionfly (= eruciform)
(Whittle and Ferguson 1987), was described by Mathur and Singh (1963)Mathur and Singh (1963):
Mathur, R. N. and P. Singh. 1963. Immature Stages of Indian Lepidoptera. No. 13 Pyralidae, Sub-family Pyraustinae. Indian Forest Records, n.s. (Entomology) 10: l17-148., Piao et al. (2008)Piao et al. (2008):
Piao, M-H., S-Z. Liu and C-Y. Lee. 2008. Descriptions of three species larvae of genus Diaphania from China. Journal Of Zhejiang Forestry College 25: 235-239., and Singh (2012)Singh (2012):
Singh, D. 2012. Morphotaxonomy of immature stages of some economically important pyraloidea (Ditrysia: Lepidoptera) of Punjabi. Punjabi University Ph.D. Dissertation. 281 pages.. Color photographs can be found in NBAII (2013)NBAII (2013):
NBAII - National Bureau of Agriculturally Important Insects. 2013. Insects in Indian Agroecosystems. Diaphania indica.. The life cycle was illustrated by Singh (2012)Singh (2012):
Singh, D. 2012. Morphotaxonomy of immature stages of some economically important pyraloidea (Ditrysia: Lepidoptera) of Punjabi. Punjabi University Ph.D. Dissertation. 281 pages..
Typically, larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of the D. hyalinata-indica complex have no genal spot, an outer tooth on the mandible, a bisetosebisetose:
two setae
SV group on A1, and abdominal crochetscrochets:
sclerotized, hooklike structures, usually arranged in rows or circles on the prolegs of Lepidoptera larvae
in a mesal penellipsemesal penellipse:
an incomplete circle of crochets closed mesally and open laterally
. The L1 setaseta:
a hairlike projection of the body wall that is articulated in a socket; compare to spine
on A8, at least in D. hyalinata, is separated from the spiracle by the vertical diameter of the spiracle (Peterson 1962: L51).
Larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of the D. hyalinata-indica complex are most likely to be confused with D. nitidalis. Unlike D. hyalinata and D. indica, D. nitidalis lacks an outer tooth on the mandible above the lateral setaeseta:
a hairlike projection of the body wall that is articulated in a socket; compare to spine
and has a genal spot (see D. nitidalis fact sheet). There are also coloration differences between the two taxa (Smith 1911Smith 1911:
Smith, R. I. 1911. Two important cantaloupe pests. North Carolina Agricultural Experimental Station Bulletin 214. 45 pp., Passoa 1985Passoa 1985:
Passoa, S. 1985. Taxonomy of the larvae and pupae of economically important Pyralidae in Honduras. Master's Thesis. University of Florida. Gainesville. 486 pp., Weisman 1986Weisman 1986:
Weisman, D. M. 1986. Keys for the identification of some frequently intercepted lepidopterous larvae. U.S. Department of Agriculture, Animal and Plant Health Inspection Service, Plant Protection and Quarantine series 81-47. 64 pp., Whittle and Ferguson 1987, Neunzig 1987Neunzig 1987:
Neunzig, H. H. 1987. Pyralidae (Pyraloidea), pp. 464-494. In F.W. Stehr (ed.). Immature Insects. Kendall Hunt Publishing Company. Dubuque, Iowa., Solis 2011Solis 2011:
Solis, M. A. 2011. Key to selected Pyraloidea (Lepidoptera) larvae intercepted at U. S. ports of entry: revision of Pyraloidea in "Keys to some frequently intercepted lepidopterous larvae" by Weisman 1986 (updated 2006).). Live larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of D. nitidalis are green, becoming copper to pink at maturity, without the white subdorsal longitudinal stripes found on D. hyalinata and D. indica. Another difference is that early instars of D. nitidalis have dark pigmented body pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
. All instars of larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
in the D. hyalinata-indica complex have pale pinaculapinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
, or at most two tiny black spots on the subdorsal pinaculumpinaculum:
a small, flat, or slightly elevated chitinized area bearing a seta or setae
of the mesothoraxmesothorax:
the second thoracic segment
and metathoraxmetathorax:
the third thoracic segment
, and do not turn pink or copper before pupation. Of course the white subdorsal lines usually fade in preserved specimens. Sometimes the mandibles of D. hyalinata show through the transparent head before molting to superficially give the impression of a genal spot which could cause the larvalarva:
the stages between the egg and pupa of those insects having complete metamorphosis
to be wrongly identified as D. nitidalis.
According to Capps (1948)Capps (1948):
Capps, H. W. 1948. Status of the pyraustid moths of the genus Leucinodes in the New World, with descriptions of new genera and species. Proceedings of the United Sates National Museum 98: 69-83., some N. elegantalis have a bisetosebisetose:
two setae
SV group on A1. These could potentially be confused with larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of the D. hyalinata-indica complex. Among many differences, the mandible of the D. hyalinata-indica complex has an outer tooth lacking in N. elegantalis.
So far no characters have been found to separate D. indica from D. hyalinata although the position of the substemmatal and stemmatal setaeseta:
a hairlike projection of the body wall that is articulated in a socket; compare to spine
may differ (Passoa 1985Passoa 1985:
Passoa, S. 1985. Taxonomy of the larvae and pupae of economically important Pyralidae in Honduras. Master's Thesis. University of Florida. Gainesville. 486 pp.). SS2 and SS3 are spaced closer than SS3 and S3 in D. indica. The distance between SS2 and SS3 is subequalsubequal:
nearly equal
to that between SS3 and S3 in D. hyalinata. The drawing in Singh (2012)Singh (2012):
Singh, D. 2012. Morphotaxonomy of immature stages of some economically important pyraloidea (Ditrysia: Lepidoptera) of Punjabi. Punjabi University Ph.D. Dissertation. 281 pages. seems to fit this diagnosis but more material is needed to evaluate the variability of this character. The larval antenna of D. indica was described in CEIR (1958), but no comparisons were made to other species. Mathur and Singh (1963)Mathur and Singh (1963):
Mathur, R. N. and P. Singh. 1963. Immature Stages of Indian Lepidoptera. No. 13 Pyralidae, Sub-family Pyraustinae. Indian Forest Records, n.s. (Entomology) 10: l17-148. used the height of the adfrontal area and position of AF1 to separate D. indica from other Diaphania species in India. This, and the pattern of pigmentation on the prothoracic and anal shields they illustrated (Mathur and Singh (1963: fig. 36, 37), potentially can help identify D. indica. Nevertheless, accurate morphological identifications are easily made by rearing because the genitalia of both sexes and pupal morphology are very different in D. indica and D. hyalinata.
Passoa (1985: figs. 173, 174) compared the pupal heads of D. indica and D. hyalinata. The pilifers of D. hyalinata are much longer than broad giving the head a long and slender appearance. The opposite is true for D. indica; the pilifers are broader than long causing the head to appear shorter and stouter. This difference is often visible on cast skins. Whittle and Ferguson (1987) added that maxillaemaxilla:
the paired mouthparts posterior to the mandibles
extend to A8 or A9 in D. hyalinata, but only to A7 in D. indica. The pupal abdominal dorsum of D. nitidalis is roughened; this separates it from D. hyalinata (Passoa 1985: figs. 170, 171) and D. indica which have punctate abdomens.
Identification of larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
in the D. hyalinata-indica complex is based on the absence of a genal spot, presence of an outer tooth on the mandible, and the bisetosebisetose:
two setae
SV group on A1. None of these characters are very distinctive. Many species of crambids lack a genal spot. The presence of an outer tooth on the mandible is unusual but a similar outer tooth is also found on Maruca vitrata, for example. The bisetosebisetose:
two setae
SV group on A1 is helpful, but again not unique. A lack of characters for recognizing the D. hyalinata-indica complex is compounded by numerous sibling species of Diaphania and relatives (in both the New and Old World) with unknown larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
. Larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of both the D. hyalinata-indica complex and D. nitidalis have a row of microspinesmicrospines:
minute spines on the body, usually visible only under magnification
at the base of the proleg but the distribution of this character in Crambidae has not been studied. These uncertainties all suggest a need for caution. Unless the specimen is clearly in the in the D. hyalinata-indica complex from the appropriate host, use Crambidae because no single character can separate the subfamilies Pyraustinae and Spilomelinae.
Origin and host are both helpful for identification of the D. hyalinata-indica complex. Diaphania indica occurs in Florida, tropical America, and the Old World tropics (Clavijo 1990Clavijo 1990:
Clavijo, J. A. 1990. Systematics of black and white species of the genus Diaphania Hubner (1818) (Lepidoptera: Pyralidae: Pyraustinae). Ph.D. Dissertation. McGill University. Montreal, Canada. 215 pp.). Diaphania hylinata occurs from Canada to Argentina (Solis 2011Solis 2011:
Solis, M. A. 2011. Key to selected Pyraloidea (Lepidoptera) larvae intercepted at U. S. ports of entry: revision of Pyraloidea in "Keys to some frequently intercepted lepidopterous larvae" by Weisman 1986 (updated 2006).). Therefore, Old World interceptions of the D. hyalinata-indica complex, especially on crops, are D. indica and not D. hyalinata.
The hosts of D. hyalinata are in the Cucurbitaceae, the one exception on Ipomea (Clavijo 1990Clavijo 1990:
Clavijo, J. A. 1990. Systematics of black and white species of the genus Diaphania Hubner (1818) (Lepidoptera: Pyralidae: Pyraustinae). Ph.D. Dissertation. McGill University. Montreal, Canada. 215 pp.) is probably it atypical. Diaphania indica has a wider host range that includes Cucurbitaceae, legumes, cotton (Clavijo 1990Clavijo 1990:
Clavijo, J. A. 1990. Systematics of black and white species of the genus Diaphania Hubner (1818) (Lepidoptera: Pyralidae: Pyraustinae). Ph.D. Dissertation. McGill University. Montreal, Canada. 215 pp.) and eggplant (CEIR 1958). Thus, interceptions of the D. hyalinata-indica complex outside of the Curcubitaceae are D. indica and not D. hyalinata. Larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
of the D. hyalinata-indica complex are most often foliage feeders, but do bore in fruit, stems and flowers, although this is not their typical biology (Clavijo 1990Clavijo 1990:
Clavijo, J. A. 1990. Systematics of black and white species of the genus Diaphania Hubner (1818) (Lepidoptera: Pyralidae: Pyraustinae). Ph.D. Dissertation. McGill University. Montreal, Canada. 215 pp.).
There is a need to clarify the relative abundance of the D. hyalinata-indica complex on Momordica at United States ports and in the New World in general, especially in Florida (Clavijo 1990Clavijo 1990:
Clavijo, J. A. 1990. Systematics of black and white species of the genus Diaphania Hubner (1818) (Lepidoptera: Pyralidae: Pyraustinae). Ph.D. Dissertation. McGill University. Montreal, Canada. 215 pp.). One D. indica was reared from a larvalarva:
the stages between the egg and pupa of those insects having complete metamorphosis
intercepted from Jamaica on Momordica (adult det. A. Solis) in 1995. Several larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
were collected at Homestead, Florida, in 1983 by S. Passoa on Momordica. These were reared to the pupal stage and identified as D. indica by pupal characters. The Florida State Collection of Arthropods has a large series of D. indica immatures from Momordica in the D. Habeck collection (for example A-725). Diaphania indica has been intercepted in consignments of Momordica from Kenya and Surinam in 2005 by the National Plant Protection Organization, the Netherlands (CSL 2005). Because of these records, it is tempting to say that D. hyalinata is very rare on Momordica and therefore D. indica is the name to use for this host. A rearing program was suggested for D. indica at U. S. ports many years ago (Passoa and Cavey 1992Passoa and Cavey 1992:
Passoa, S. and J. F. Cavey. 1992. D. indica-hyalinata complex. Potential target fact sheet number 5. Northeastern Exotic Pest Survey Committee unpublished aid. 2 pp.). It is only necessary to rear larvaelarva:
the stages between the egg and pupa of those insects having complete metamorphosis
to the pupal stage. Of course, modern methods like DNA barcoding would allow for more rapid identification.
Members of the Diaphania hyalinata-indica complex have been intercepted from the following locations:
Antigua and Barbuda, Barbados, Cayman Islands, China, Colombia, Costa Rica, Cuba, Dominica, Dominican Republic, Ecuador, El Salvador, Ghana, Grenada, Guatemala, Guyana, Haiti, Honduras, India, Israel, Jamaica, Liberia, Mexico, Morocco, Nigeria, Palestinian Territory, Puerto Rico, Somalia, St. Lucia, St. Maarten, Trinidad and Tobago, Turks and Caicos Islands, US Virgin Islands
Members of the Diaphania hyalinata-indica complex have been intercepted on the following hosts:
Abelmoschus esculentus, Abelmoschus sp., Alstroemeria sp., Amaranthus sp., Annona muricata*, Antirrhinum sp., Apium graveolens, Artocarpus altilis, Cajanus cajan, Capsicum frutescens, Carica papaya, Cercis sp., Chenopodium berlandieri ssp nuttalliae, Chenopodium sp., Chrysanthemum sp., Citrus sp.*, Coccinia grandis, Coccinia sp., Coccoloba uvifera*, Cocos sp., Corchorus sp., Coriandrum sativum, Cucumis sativus, Cucumis sp., Cucurbita maxima, Cucurbita moschata, Cucurbita pepo, Cucurbita sp., Cucurbitaceae, Curcubita sp., Cymbopogon sp., Jatropha sp., Lagenaria siceraria, Limonium sp., Luffa acutangula, Luffa aegyptiaca, Luffa sp., Malvaceae, Mentha sp., Momordica balsamina, Momordica charantia, Momordica indica, Momordica sp., Moringa citrifolia, Opuntia sp.*, Origanum sp., Pachyrhizus erosus, Petroselinum crispum, Phaseolus sp., Physalis pubescens, Physalis sp., Phytolacca sp., Piper sp., Porophyllum ruderale, Portulaca oleracea, Salvia officinalis, Sechium edule, Sechium sp., Senna sp., Smilax sp., Solanum melongena*, Solanum sp.*, Solanum torvum*, Stachytarpheta jamaicensis, Thymus caespititius, Thymus vulgaris, Trichosanthes sp., Xanthosoma brasiliense, Zea mays*
Although D. indica has a wide host range, records on Annona, citrus, seagrape, Opuntia, Solanum, and corn are among those needing confirmation (indicated here with an *). The normal hosts of D. indica are in the Cucurbitaceae, Leguminosae, and Malvaceae (CSL 2005).
Click here to download a full-size printable PDF of this larval setal map
Click here to download a PDF of this fact sheet content and images